正常分化的细胞主要依靠线粒体的氧化磷酸化为细胞供能,而大多数肿瘤细胞则依赖有氧糖酵解,这种现象被称为 「Warburg effect」 。 有氧糖酵解产生 ATP 的效率很低,但却赋予肿瘤细胞很多优势。
2009 年,Lewis C. Cantley 和 Craig B. Thompson 等人在【Science】上」Understanding the Warburg Effect: The Metabolic Requirements of Cell Proliferation「的文章系统综述了「Warburg effect」。
肿瘤细胞以及所有增殖细胞的代谢都会促进营养物质的摄取和吸收,以合成新细胞所需的生物质(biomass)(如核苷酸、氨基酸和脂质)。最近的一些研究表明:(Ⅰ)与细胞增殖有关的几条信号传导通路也可以调节代谢通路,负责将营养物质合成生物质;(Ⅱ)某些与癌症相关的突变使肿瘤细胞能够以有利于增殖而不是高效产生ATP的方式获取和代谢营养物质。 更好地了解细胞代谢与生长控制之间的联系最终可能有利于对癌症的治疗。
对于单细胞生物,比如微生物,存在着进化压力,这就要求它们在营养物质充足时必须尽快繁殖。 它们的新陈代谢控制系统已经进化,当感应到营养物质充足时,就将碳、氮和 ATP 作为新细胞所需的原料。当营养缺乏时,细胞会停止合成生物量并适应新陈代谢,最大化地产生 ATP 来生存(图1)。

在多细胞生物中,大多数细胞的营养是源源不断的。机体存在控制系统,当营养供应超过细胞分裂所需的水平时,该系统可以防止单个细胞的异常增殖。哺乳动物细胞通常不会从周围环境吸收营养,除非受到生长因子的刺激,所以会阻止不受控制的增殖。肿瘤细胞通过基因突变从功能上改变受体启动信号通路,克服了对生长因子的依赖。
越来越多的证据表明,其中某些途径可组成性激活营养吸收和代谢,从而促进细胞存活和生长。癌基因的突变会促进营养物质摄取,尤其是葡萄糖,从而满足或超过细胞生长和增殖的生物能需求。 这种认识使人们想起了 Otto Warburg 在 1924 年的发现,即肿瘤细胞与正常组织细胞以不同的方式代谢葡萄糖。通过研究 Louis Pasteur 关于如何在哺乳动物组织实现葡萄糖乙醇发酵的观察, Warburg 发现,与大多数正常组织不同,肿瘤细胞即使在氧气足以支持线粒体氧化磷酸化的情况下,也倾向于将葡萄糖「发酵」成乳酸。 Warburg 的观察仍缺乏确切的解释,至少乍看之下,线粒体氧化磷酸化完全分解葡萄糖可以最大程度地满足细胞增殖所需的 ATP。
增殖的哺乳动物细胞存在合成代谢
我们目前对代谢途径的理解主要是基于分化组织中非增殖细胞的研究。 在有氧条件下,大多数分化细胞主要通过线粒体三羧酸(TCA)循环将糖酵解产生的丙酮酸氧化为二氧化碳。该反应产生 NADH,然后促进氧化磷酸化,最大限度地合成 ATP,而乳酸最少 (图2)。只有在厌氧条件下,分化细胞才会产生大量的乳酸。 相反,大多数肿瘤细胞会产生大量的乳酸,而与氧气存在与否无关,因此,肿瘤细胞的代谢通常被称为「有氧糖酵解」。

Warburg 最初猜测肿瘤细胞的线粒体存在发育缺陷,从而导致有氧呼吸功能受损,故而依赖于糖酵解代谢。但是,随后的研究表明,大多数肿瘤细胞中线粒体功能并未受损,表明肿瘤细胞的有氧糖酵解存在其他解释。
为什么增殖细胞会选择效率较低的代谢方式?
如上所述,许多单细胞生物通过发酵(有氧糖酵解的微生物当量)进行增殖,与人的肿瘤细胞类似,即使在氧气充足的情况下,也优先发酵葡萄糖(图1)。这表明有氧糖酵解代谢可以为细胞增殖提供足够的能量。 葡萄糖代谢为乳酸时,每个葡萄糖分子仅生成 2 个 ATP,而一个葡萄糖分子完全氧化后,氧化磷酸化最多生成 36 个 ATP。
这就提出了一个问题,为什么在增殖细胞中会选择 ATP 产生效率较低的代谢方式?一种可能的解释是,仅当营养缺乏时,ATP 产生效率低才是问题。 增殖的哺乳动物细胞并非如此,哺乳动物细胞暴露在循环血液中,有持续供应的葡萄糖和其他营养物质。直到最近增殖细胞的代谢途径及其调控才开始研究,并且有证据表明,ATP 永远不可能是限制因素。无论刺激多少,有氧糖酵解的细胞其 ATP/ADP 和 NADH/NAD+比值都很高。此外,即使是 ATP/ADP 比例的微小变化也会影响其生长。ATP 不足的细胞通常会发生凋亡。当正常的增殖细胞葡萄糖代谢产生 ATP 的能力受损时,它们会经历细胞周期停滞并重激活分解代谢,同时存在信号通路来感知能量状态。其中最显著的特征是腺苷酸激酶的活性,该酶通过将两分子 ADP 转化为一分子 ATP 和一分子 AMP 来缓冲 ATP 的减少。随着 ATP 的减少,这有助于维持 ATP/ADP 比例,但是 AMP 的积累会激活 AMPK。 这种激活取决于肿瘤抑制蛋白 LKB1,进而导致多个靶蛋白磷酸化,从而改善细胞中的能量电荷。LKB1 最初被认为是一种抑癌基因,这表明在某些细胞类型中,感知能量压力的能力可能是阻止恶化的一个重要检查点。
第二种可能的解释是,增殖细胞之所以选择有氧糖酵解,是有重要的代谢需求,超出了 ATP 的范畴。
从数字上看,增殖细胞的代谢需求是什么?
有丝分裂产生两个子代细胞,增殖细胞必须要复制其所有的细胞内含物,包括核苷酸、氨基酸和脂质。在生长过程中,葡萄糖被用于合成这些物质(biomass)以及产生ATP。尽管ATP水解为生物质合成的某些生化反应提供了自由能,但这些反应还有其他需求。
例如,棕榈酸酯——细胞膜的主要组成部分,它的合成需要7分子ATP,8分子乙酰CoA(16分子碳)和14分子NADPH(28分子电子)。同样,氨基酸和核苷酸合成消耗的碳和NADPH当量比ATP要多。 1分子葡萄糖最多可以生成36分子ATP或30分子ATP和2分子NADPH(磷酸戊糖途径),或者为大分子合成提供6个碳。 因此,要形成16个碳原子的脂肪酰基链,单个葡萄糖分子可以提供5倍的ATP,但是需要7分子葡萄糖才能满足所需的NADPH。这种35倍的不对称性只能通过消耗3分子葡萄糖生成乙酰CoA来部分补偿,从而满足酰基链碳的需求。
不言而喻,对于细胞增殖,大部分葡萄糖不能用来产生ATP。否则ATP / ADP比率增加将严重减少糖酵解中间体通量,从而限制大分子合成所需的乙酰CoA和NADPH。
对于培养的大多数哺乳动物细胞来说,仅有的能量来源是葡萄糖和谷氨酰胺。这意味着葡萄糖和谷氨酰胺将提供细胞生长和分裂所需的大部分的碳、氮、自由能以及还原当量。 从这个角度来看,很明显,线粒体氧化磷酸化将所有葡萄糖转化为 CO2 以最大化产生 ATP,与增殖细胞的需求背道而驰。 葡萄糖可以转化为合成大分子的前体,如用于脂肪酸合成的乙酰 CoA、非必需氨基酸合成的糖酵解中间体以及核苷酸合成的核糖。 这至少部分解释了 Warburg 效应。
最近的 13C-核磁共振波谱检测表明,培养中的胶质母细胞瘤细胞将其 90%的葡萄糖和 60%的谷氨酰胺转化为乳酸或丙氨酸。 尽管大多数乳酸和丙氨酸是作为废物从细胞中排出的,但乳酸和丙氨酸的生成伴随着大量 NADPH 的产生(图3)。 除了通过转氨反应生成非必需氨基酸外,谷氨酰胺分解为乳酸的过程中通过苹果酸脱氢酶产生 NADPH。生长因子信号转导也会调节丙酮酸激酶的活性,并通过糖酵解的后续步骤调节碳通量。对丙酮酸激酶的调节可以促进葡萄糖代谢物重新定向到磷酸戊糖途径,以及核苷酸和氨基酸的生物合成途径。葡萄糖和谷氨酰胺向乳酸的转化都涉及到乳酸脱氢酶(LDH),抑制 LDH 活性会干扰细胞排泄多余碳源的能力,从而影响细胞增殖。而消除多余碳源又是必须的,唯有如此才能产生足够的 NADPH 维持细胞增殖。
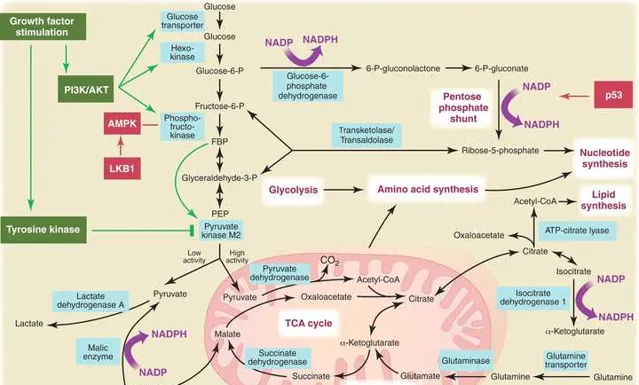
用于脂肪酸合成的大部分碳来自葡萄糖。在此过程中,葡萄糖首先在线粒体基质中转化为乙酰 CoA,进而通过 TCA 循环转化为柠檬酸。在大多数增殖细胞中,ATP/ADP 和 NADH/NAD+的比率高,柠檬酸会从线粒体排出到胞质中用于合成脂肪酸。在胞质中,柠檬酸重新转化为乙酰 CoA,作为酰基链的碳源。 由柠檬酸合成乙酰 CoA 需要酶 ATP-柠檬酸裂解酶(ACL),破坏 ACL 会影响肿瘤生长。 谷氨酰胺的摄取对脂质合成也很关键,它以线粒体草酰乙酸的形式提供碳,维持 TCA 循环第一步中柠檬酸的产生。 因此,协调谷氨酰胺和葡萄糖代谢,以维持脂肪酸合成所需的乙酰 CoA 和 NADPH。
伴随 Warburg 效应产生的过量乳酸似乎是细胞资源的低效利用。每从细胞排出一分子乳酸就会浪费三个碳,这些碳原本可以用于生成 ATP 或大分子合成的前体。可能将多余的碳以乳酸形式排出是有效的,因为它可以使碳更快地结合到生物质中,从而促进细胞快速分裂。对于大多数增殖细胞,营养物质没有限制,因此没有选择压力来优化代谢以提高 ATP 产量。
相反,确实存在代谢速率的选择压力。免疫反应和伤口修复取决于效应细胞增殖扩散的速度。为了生存,机体必须向反应细胞发出信号,以使其合成代谢生长速率最大化。能将葡萄糖和谷氨酰胺有效转化为生物质的细胞增殖最快。 对于机体而言,营养可能出现缺乏,在特定的、非增殖组织中有其他途径可以回收增殖细胞快速生长过程中丢弃的过量乳酸和丙氨酸。 肝脏中的 Cori 循环可以将活跃的增殖组织所产生的乳酸再循环为葡萄糖,并且有类似的途径再循环谷氨酰胺「无效」代谢所产生的丙氨酸。在免疫应答或伤口修复过程中通过再循环回收细胞增殖所产生的有机废物,对机体的能量储备影响很小。此外,新出现的证据表明,肿瘤细胞代谢可能是异质的,有些细胞可以利用产生的过量乳酸作为线粒体氧化磷酸化的原料。
参考文献:Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Science . 2009;324(5930):1029‐1033. doi:10.1126/science.1160809











