最近写了一篇长文【关于维生素B12全合成的详细解析】,就来这里投一下。
相信看完之后,你就明白全合成的意义了。
------------
谨以此文,纪念维生素B12被人类发现90周年。
注:如果本文中有任何错误或疏漏,请务必联系作者或在评论中写出。不胜感谢!
1.前情提要
维生素B12 (\text{Vitamin B12} ,又名 钴胺素 ,缩写 \text{VB12} ),这种物质(其实严格来说是一类物质)如今早已家喻户晓。作为唯一一种 含有金属元素的维生素 ,其中心的 \ce{Co} 原子赋予了它鲜红的色彩,也让它有了一个别称: 红色维生素 。

巧合的是,鲜红的维生素B12最重要的作用之一,就是为我们的身体 制造鲜红的血液 。具体来说,维生素B12是一种 辅酶 ,参与红细胞的制造与成熟,也参与某些氨基酸(如 蛋氨酸 )和 胸腺嘧啶 在体内的合成。总之,它在人体内发挥着非常重要的作用,是 维持正常代谢和机能不可或缺的物质 。
同时,维生素B12是也 结构最复杂的维生素 。它的化学式为 \ce{C63H88N14O14PCo} ,分子量高达 1355.37 ,拥有一个 咕啉 (corrin )大环(注意它与 卟啉 结构的区别)、一条结构类似于 核苷酸 的长链、14 个具有手性的碳原子和一个在中心配位的 钴原子 ,如下图:
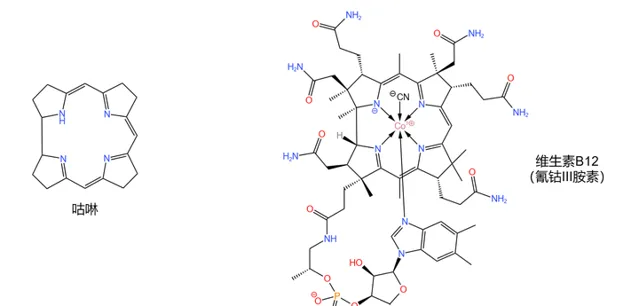
(注:严格来说, 氰钴 \text{(III)} 胺素 只能算是维生素B12的一种,但一般可以把它当做维生素B12的代表。下文中所有「维生素B12」字眼基本都指代这一物质。)
虽然维生素B12对正常生理活动非常重要,但是高等动植物却 不能自行制造维生素B12 。自然界中,这种物质主要依靠 细菌 等微生物供给,而我们则必须依靠摄食来补充。
人类对维生素B12的认识是 漫长而曲折 的。19 世纪 50 年代,英国医生 Addison 首次描述了一种致死的 恶性贫血 ,并发现这种疾病与患者的 胃部受损 有关。之后的几十年间,人们一直在试图搞清楚这种病的根源,但进展寥寥。直到 1929 年,美国科学家 Castle 提出了著名的 双因子理论 来解释发病机制:病人由于胃部受损,缺乏一种 内因子 (intrinsic~factor ),导致其无法从食物中吸收一种 外因子 (extrinsic~factor ),进而导致患病。
刚开始,许多科学家都对这套理论不以为然。直到 1934 年,G.~Whipple 、G.~Minot 和 W.~Murphy 三人真的发现且分离出了这种「外因子」,并将其取名为 维生素B12 ,对应的争论才偃旗息鼓。三人也因为这项重大成果获得了诺贝尔生理学或医学奖:

(上图:三位发现维生素B12的科学家)
1948 年,Rickers~E.~L. 等人得到了 纯的维生素B12 。随后,对其复杂结构的破译轰轰烈烈地展开。8 年之后,1956 年,D.~Hodgkin 等人用 X射线衍射 法彻底破解了它的结构,并因此获得了 1964 年的诺贝尔化学奖。
好,既然结构已经确认,那么下一步当然就是合成这种物质。可是,它的结构是 如此巨大而复杂 ,简直令人望而生畏。当时的化学家们都将其称为 有机合成界的珠穆朗玛峰 。(虽然后面又出现了更加复杂的物质(Link:岩沙海葵毒素的全合成)拿走了这一称谓)
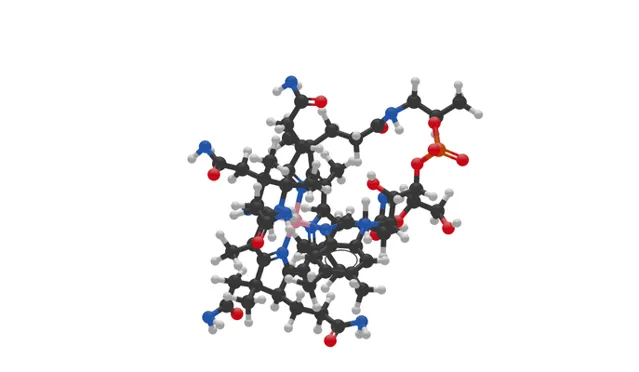
(上图:维生素B12的球棍模型,其中黑色:C ,白色:H ,红色:O ,橙色:P ,蓝色:N ,粉色:Co )
但是,人类并不会因此放弃。
从 1959 年开始,瑞士化学家 A.~Eschenmoser 等人就开始致力于攀登这座高峰,成为了这项工作的 先行者 。1960 年,德国化学家 Bernhauer~K. 等人加入了攀登者的行列,并成功完成了从 钴啉胺酸 (cobyric~acid )到维生素B12的 半合成 。1961 年,刚刚完成 叶绿素A 全合成(Link:叶绿素A的全合成)的「 有机合成艺术家 」R.~Woodward 带领着他的课题组加入了进来,并很快与其他研究者取得合作。最终,1972 年,在整整 13 年的光阴之后,这项超过 100 位研究者参与的、跨越半个地球的「 有机合成化学史上最疯狂的工作 」终于宣告成功。

(上图:【维生素B12的全合成】论文影,1973 ,伍德沃德。参与工作的其他课题组也发表了各自的论文。)
在下文中,我们将会详细讨论这条凝聚了上百人心血的维生素B12全合成路线。 不要因为它的复杂而感到畏惧 ,请相信我,当你逐渐理解它的时候,你会发现它的精巧和美丽。它是一件巧夺天工的艺术品。
在上帝创造的自然界的旁边,化学家们又创造了一个新的世界。 ——伍德沃德,19562.概述
与叶绿素A一样,维生素B12也是一个 相当脆弱的分子 。它只有在 \text{PH}=4.5-5.0 的狭小区间内才能长时间稳定,而一旦 \text{PH}>7 或是 \text{PH}<2 就会迅速分解。它的热稳定性也不好,在 100^{\circ}C 以上也会彻底分解。这无疑是给全合成工作带来了巨大的挑战。
但是,这还并不是合成过程中最大的难点。前面提到,维生素B12拥有一个 咕啉大环 (下图中紫色部分)——这是整个分子的 核心结构 。它由 4 个 氢化的吡咯环 拼接而成,具有显著的 不对称性 。由于无法形成 \pi 电子的环流,咕啉环 没有芳香性 ,性质十分脆弱,且周边分布着众多的 手性碳原子 。当时的手性合成技术还远没有现在成熟,因此在维生素B12之前,人们还从来没有成功合成过咕啉衍生物。
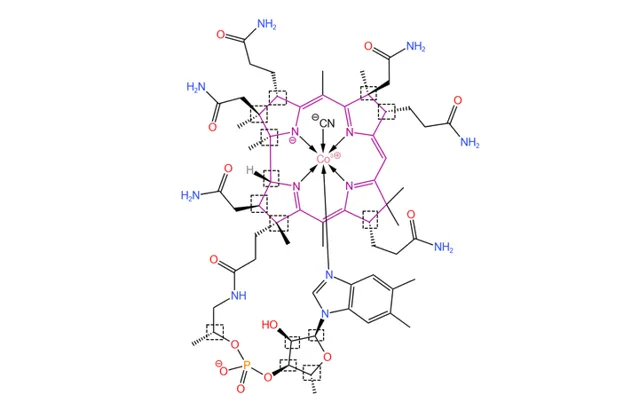
(上图中:紫色标出了咕啉结构;虚线框框出了所有手性碳原子)
1960 年,分子下方的那条类似于核苷酸的长链被 Bernhauer~K. 等人成功合成——这也算是全合成中的 第一个阶段性成果 。于是,后续合成工作的目标变成了 钴啉胺酸 (如下图),而咕啉结构的构建也随即成为了核心问题:
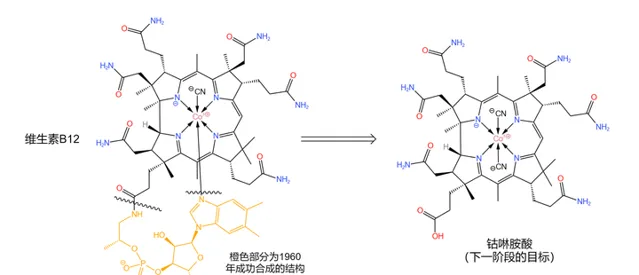
解决这一问题的思路是 将整个分子劈成两半 (如下图),然后再将它们拼接到一起。其中,左半部分被命名为 cyanobromide ,由 R.~Woodward 的团队负责合成;右半部分被命名为 thiodextrolin ,由 A.~Eschenmoser 的团队负责合成:
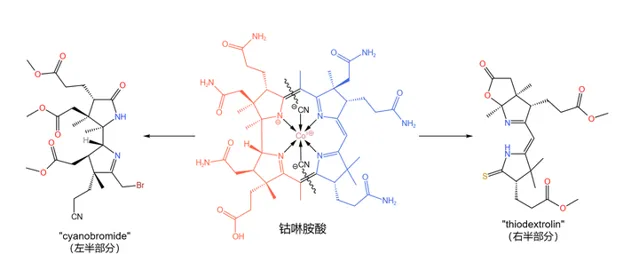
现在,分工已经确定,全合成的介绍也将正式开始。
本文中涉及的主要有机反应清单:
- Kucherov 加成;
- Wittig 反应;
- Birch 还原;
- Criegee 臭氧化反应;
- 羟醛缩合反应;
- Beckmann 重排;
- Diels-Alder 反应;
- Arndt-Eistert 同系增碳反应;
- Eschenmoser 偶联反应;
- Eschenmoser 酰胺水解反应;
- Meerwin 盐的烷基化反应。
3.逐步合成分析
3.1 左半部分的合成
左半部分的合成起始于一种普通的 吲哚 衍生物——6- 甲氧基-2,3- 甲基吲哚——虽然现在它看上去似乎与目标完全不沾边。首先,用 格式试剂 (这里使用的是 甲基碘化镁 ,\ce{MeMgI} )与 溴丙炔 与它反应,在 3 号位上接上一个 炔丙基 :
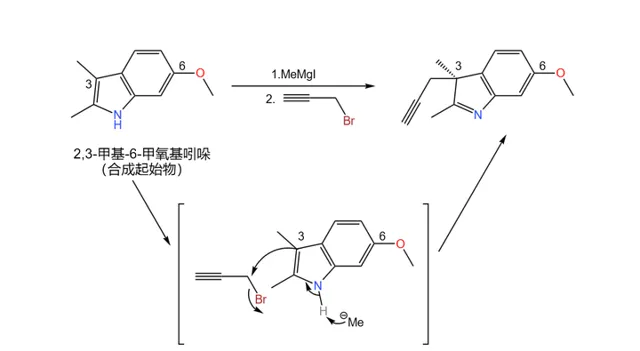
实际上,这个反应的立体选择性并不好,得到的产物是 外消旋体 ,在后续的步骤中会将其分离。
下一步,在 路易斯酸 (此处使用 \ce{BF3} )与 \ce{Hg(II)} 物种的催化下,碳碳叁键与甲醇发生著名的 Kucherov 加成反应,得到 烯基醚 ;随后,分子内自发环化,最终得到一个三环结构的酮:
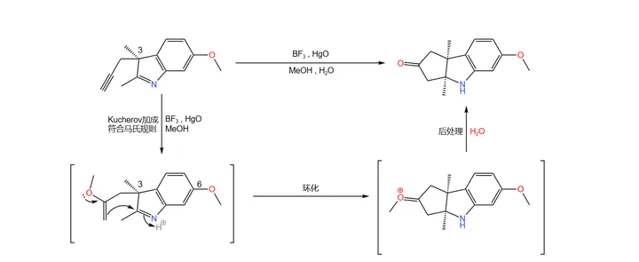
(注:由于 3 号位有甲基的存在,导致 有一侧的进攻受阻 ,因此产物的立体构型是单一的。)
现在,我们先把产物放在一边备用,来处理第二个合成起始物。它被称作 樟烯酸甲酯 (methyl~laurolenate ,见下图),是 樟脑 经过一系列反应后得到的产物,很早之前就被人们合成。伍德沃德之所以选择它,主要是因为它拥有一个继承自樟脑的手性碳原子。
首先,用 氢化铝锂 (\ce{LiAlp} )处理 \text{(S)}- 樟烯酸甲酯,将其还原为醇;随后再用 铬酸 (\ce{pCrO4} )氧化,得到一种醛;最后,使用著名的 Wittig 反应得到一个碳碳双键:
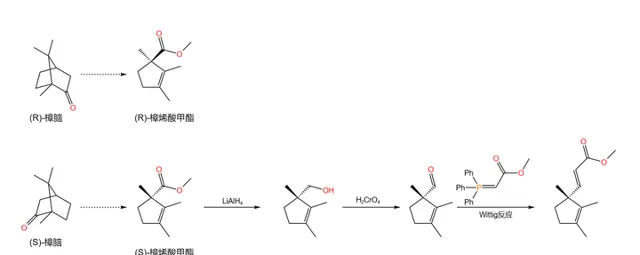
Wittig 反应是一个重要的烯烃合成反应,其机理在之前的几篇文章中都有提及,在此便不再赘述。
接着,我们用酸将得到的酯水解,再用 氯化亚砜 (\ce{SOCl2} )将其变为 酰氯 。酰氯随即与刚才合成的那个三环结构的酮反应,两者结合在一起形成酰胺结构:
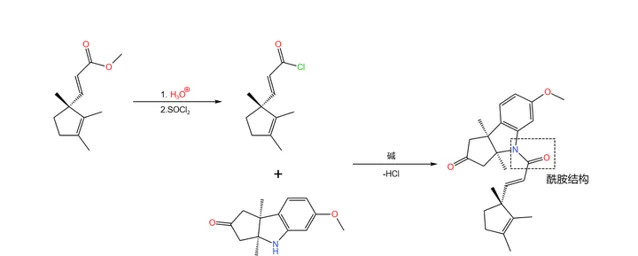
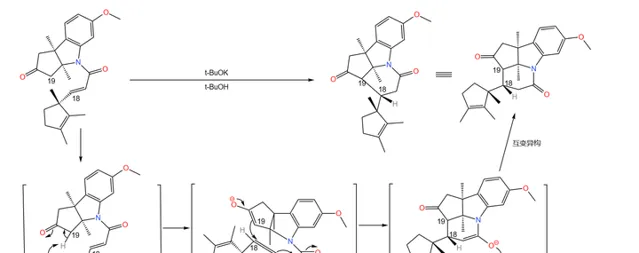
随后是一个精彩的环化反应:在 丁醇钾 (\ce{t-BuOK} )的作用下,19 号碳原子上的氢原子被夺去,形成 烯醇负离子 ;烯醇负离子进攻附近 \alpha,\beta- 不饱和的羰基环化。由于 分子的构型已经固定 ,进攻只有一个方向,因此得到了单一立体构型的产物:
(注:这里的编号表示了该碳原子最终在咕啉环中的位置)
接下来,用 乙二醇 (\ce{HO(Cp)2OH} )保护 2b 号碳原子上的酮羰基。由于分子中还有一个酰胺结构(位于 18b 号碳原子),所以我们还得用 Meerwin 盐再次进行保护(注:酰胺中的羰基无法被乙二醇保护):
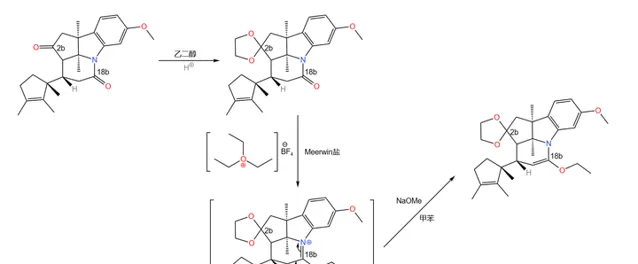
之所以要这样做,是因为下一步要用到一个比较强烈的反应,即著名的 Birch 还原反应。在 液氨 中,用过量的碱金属(这里使用 Li )处理这一物质,右上方的苯环被还原为 环己二烯 结构;随后用酸处理之,经过 双键移位 后得到 \alpha,\beta- 不饱和酮;同时,上一步的保护也被脱去:
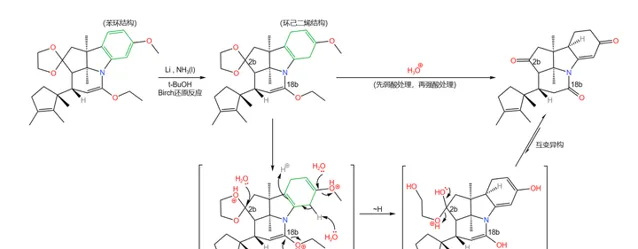
Birch 还原是非常经典的 自由基还原反应 ,在有机化学史上具有重要的地位。反应机理如下图所示:
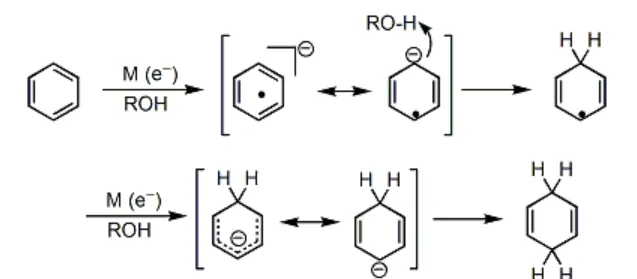
显然,对于带有 取代基 的苯环来说,最终生成的烯烃的位置选择性,是由 中间体的稳定性 决定的。一般来说,连接着 吸电子基团 (\text{EWG} )的碳原子最终不会带有双键,而连接着 给电子基团 (\text{EDG} )的碳原子最终会带有双键。例如:
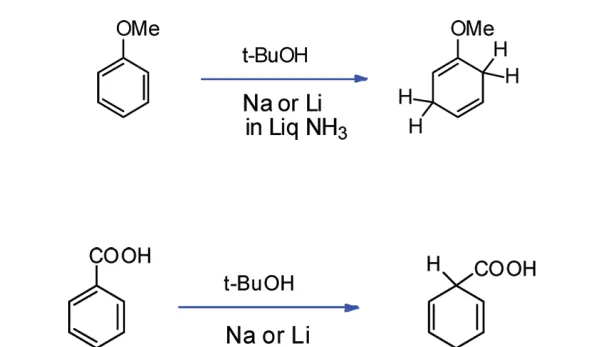
(注:想要更深入了解 Birch 还原的位置选择性的读者请见参考文献 [11] )
如果具体到我们这个反应,你会发现:苯环的 a,b,d 三个位置都带有给电子基团,所以 还原产物是单一的 。没错,这也是伍德沃德刻意的设计:
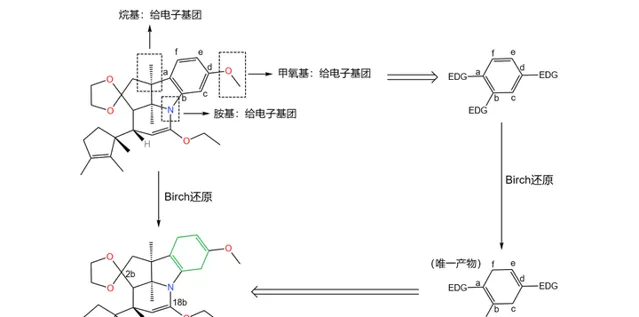
应该说,课题组的设计总体来说是成功的——两步反应的总产率达到了惊人的 90\% 。但它们很快发现了一个问题:产物中 3 号碳原子的 构型与预料中的不符 。
理论上,从 化学动力学 角度,由于附近甲基的 空间位阻 ,\ce{H} 原子应该结合在甲基的异侧;但从 化学热力学 角度,\ce{H} 原子与甲基在同侧时能量更低,也就更加稳定。但实际是,我们得到的 几乎全是热力学控制的产物 ——而那并不是我们想要的。课题组尝试改变反应条件,但都无济于事:
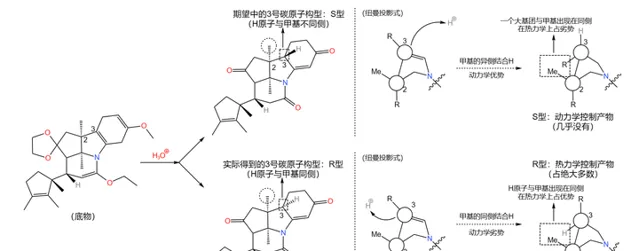
在关键时刻,伍德沃德表现出了超乎常人的冷静。经过分析,它认为:3 号碳原子是特殊的,在特定条件(比如酸性或碱性条件)下,它可以通过 互变异构 的方式转换自己的构型(注:一般来说,构型固定之后不会轻易翻转,因为 构型翻转需要断开化学键 )。由于 物质总是自发地趋向于稳定 ,所以并不是我们制不出 S 型,而是 S 型产物会迅速变成在热力学上更稳定的 R 型:
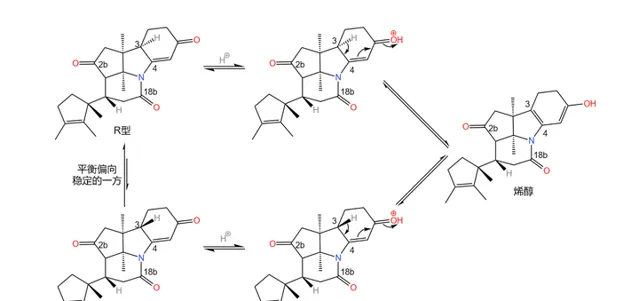
既然如此,我们是不是也可以利用这一特性,把构型翻转回来?只需要通过某种手段,让 S 型比 R 型在热力学上更稳定就行了。于是,他在论文中写道:
我们相信,如果他能在我们不需要的时候大量生成,那么至少在原理上,对它进行必要的翻转不是一件困难的事情。 ——【在天然产物全合成中的最新进展】,伍德沃德,1968他决定,暂时先不管这个构型问题,继续原有的计划。
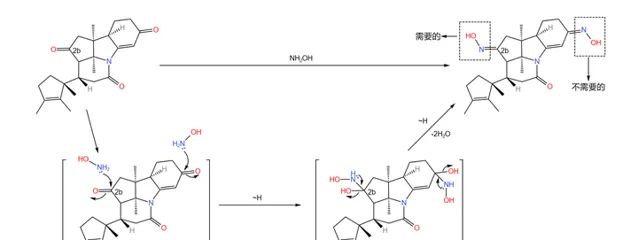
下一步,用 羟胺 (\ce{NpOH} )处理产物,把分子中的两个羰基都变成 肟 。事实上,只有 2b 号碳原子上的肟是我们需要的,但是对于羟胺来说,两个羰基的活性差不多(即使其中一个羰基是 \alpha,\beta- 不饱和的),因此我们很难在这一步区分二者,只好先把它们全部变成肟:
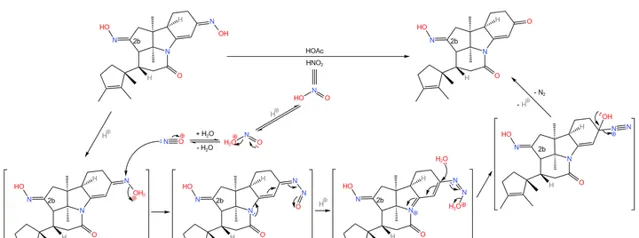
当然,我们还要把那个不需要的去掉。在非常温和的条件下,用一定量的 亚硝酸 (\ce{HNO2} )与乙酸处理,可以把不需要的那个 \alpha,\beta- 不饱和的肟基除掉,而 2b 号碳原子上的肟基不受影响:
事实上,2b 号碳原子上的肟基并不会马上用到,但后续的反应会产生越来越多的羰基,如果把这一步放到后面,那反应将 更加难以控制 。读者可以先猜一猜这个肟基后续会发挥什么作用。
回归正题,接下来登场的是著名的 Criegee 臭氧化反应。在低温下用臭氧处理该物质,分子内的两个碳碳双键被打开,形成 臭氧化物 中间体;随后,加入 高碘酸 (\ce{pIO6} )氧化;最后,加入 重氮甲烷 (\ce{CpN2} )进行酯化:
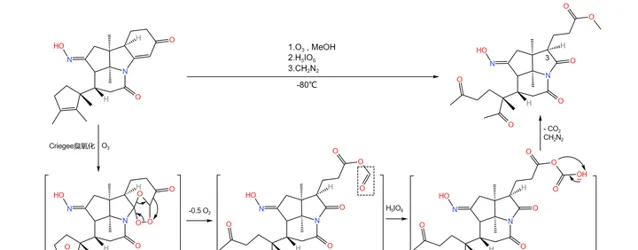
生成 臭氧化物中间体 的机理如下图所示。由于这一中间体很不稳定,所以 Criegee 臭氧化一般需要在低温下进行。
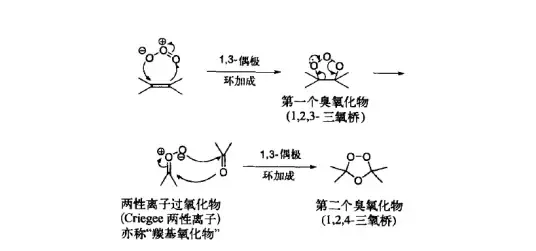
接下来,在 四氢吡咯 的催化下,分子左下角发生经典的 羟醛缩合反应 ,得到 \alpha,\beta- 不饱和酮的结构;随后,用 甲磺酰氯 (\ce{MsCl} )处理,将 2b 号位上的肟基保护起来;最后,再来一遍 Criegee 臭氧化,双键断裂,得到 17c 号位的酯结构:
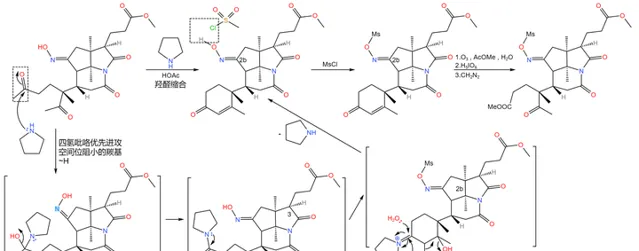
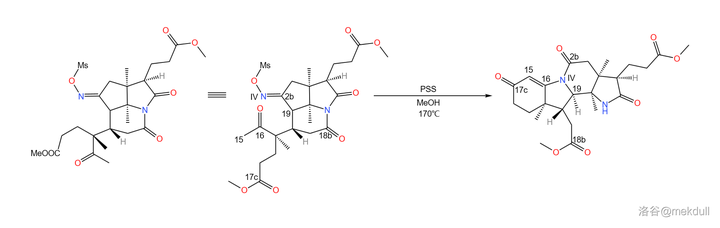
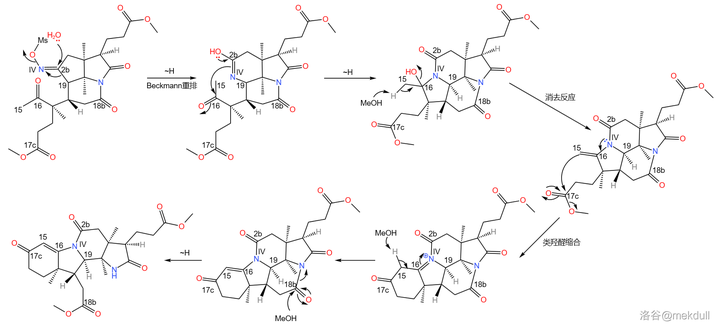
之所以如此大费周章,是因为伍德沃德准备要下一局大棋。现在,2b 号位上的肟基终于真正要出场了——它将引导著名的 Beckmann 重排反应。在 170^{\circ}C 下,用 聚苯乙烯磺酸 (\ce{(C8H8O3S)_{n}} ,缩写 \ce{PSS} )和甲醇处理之,首先发生碳骨架重排,19 号碳原子与 2b 号碳原子分离;随后,\ce{IV} 号氮原子 亲核进攻 附近 16 号碳原子的羰基,形成一个 五元环 ;接着,中间体消去 1 分子水,形成 15,16 号碳原子上的双键,从而形成 烯胺 结构;最后,烯胺 亲核进攻 附近 17c 号碳原子上的酯基,在类似于羟醛缩合的反应后形成一个六元环状的 \alpha,\beta- 不饱和酮结构。与此同时,在 18b 号位上,原来的内酰胺环被打开,从而完成整个反应历程:
或许你已经在感叹这个反应的巧妙,但相信我,精彩并未结束。我们得到的产物被命名为 半咕降甾酮 (corrnorsterone ,关于这个名字有兴趣的可以看下面的「说文解字」)。还记得 3 号位那个错误的构型吗?课题组发现,在一系列反应后,居然有 一小部分产物的构型又翻了回来 。也就是说,我们实际得到的是两种 对映异构体 的混合物。为了便于区分,课题组在它们的名字前加上了 \alpha- 和 \beta- 的前缀,如下图:
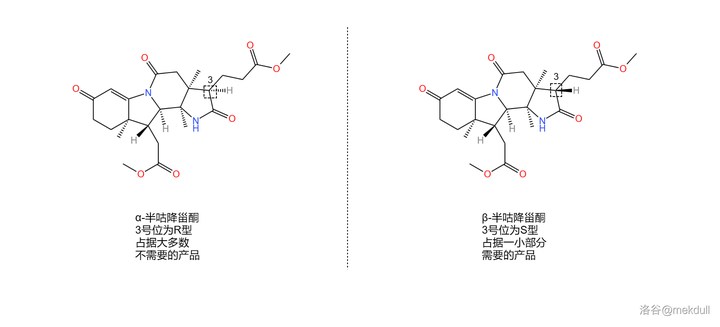 说文解字
:\text{corr-norster-one}
(半咕降甾酮) 「corr
」是单词 corrin
(咕啉)的一半,表示这种物质最终会变成咕啉环的左半部分; 「norster
」源自单词 norsteroid
(
降甾醇
,也泛指
去甲甾族化合物
),表示如果把 N
原子换成 C
原子的话,那么这个物质就具有降甾醇的碳骨架; 「one
」是酮类化合物的后缀,源自单词 ketone
。
说文解字
:\text{corr-norster-one}
(半咕降甾酮) 「corr
」是单词 corrin
(咕啉)的一半,表示这种物质最终会变成咕啉环的左半部分; 「norster
」源自单词 norsteroid
(
降甾醇
,也泛指
去甲甾族化合物
),表示如果把 N
原子换成 C
原子的话,那么这个物质就具有降甾醇的碳骨架; 「one
」是酮类化合物的后缀,源自单词 ketone
。
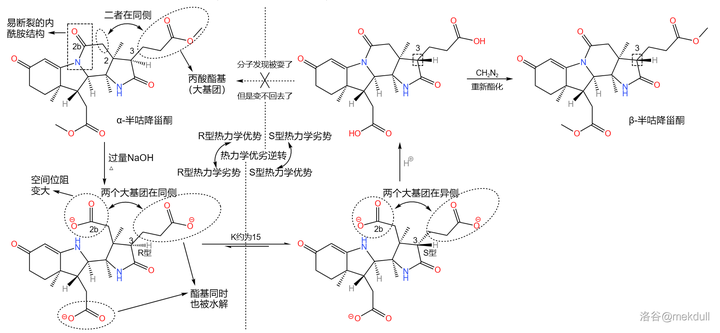
虽然构型翻转回来的只占了一小部分,但这给了课题组解决问题的希望。他们很快又发现,2b 号位上的 内酰胺 结构可以在强碱的作用下断裂。于是,一个精妙的翻转构型的方法出现了。
观察 2 号碳原子,你会发现它左侧的 2b 号碳原子因为内酰胺结构而 被「锁」在了一个固定的位置 ,连带着附近的 2a 号碳原子也无法自由移动。这就相当于,2 号碳原子的左侧是一个「 动不了 」的基团,所以空间位阻自然也比较小。而一旦我们用强碱把内酰胺环打开,那么这个基团就会重新「获得自由」, 产生巨大的空间位阻 。
那这又有什么用呢?前面提到,3 号碳原子是特殊的,在合适条件(比如酸性或碱性条件)下,它的构型可以 通过互变异构翻转 。我们观察到,在 \alpha- 半咕降甾酮中,3 号碳原子右侧的 丙酸酯基 与 2 号碳原子左侧的基团是 位于同侧 的。原来在被「锁住」时,由于空间位阻小,这种构型在热力学上是有优势的,但现在那个 被锁住的已经「解放」了 ,空间位阻急剧增大,这种构型就变成了两个大基团贴在一起, 热力学上的优劣就此逆转 。于是,3 号碳原子的构型翻转,从 R 型变成了 S 型。最后,我们只需要重新加酸至中性,分子就会重新环化——此时就算分子发现自己被耍了,构型也 变不回去 了,因为碱性条件已经丧失。就这样,我们成功把 \alpha- 半咕降甾酮变成了需要的 \beta- 半咕降甾酮:
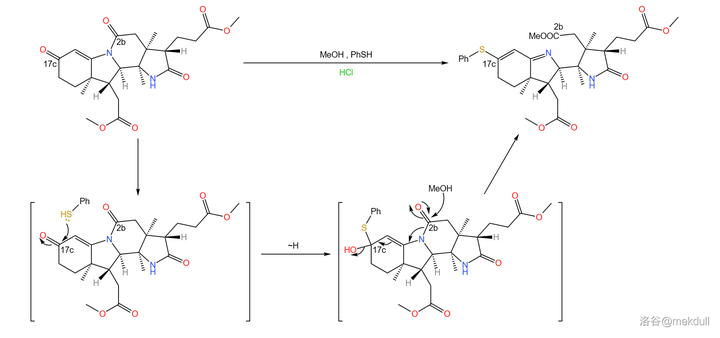
接下来,在酸性条件下,用甲醇和 苯硫醇 (\ce{PhSH} )的混合物处理 \beta- 半咕降甾酮。由于活性差别,二者会分别进攻 2b 号位和 17c 号位上的羰基。最终,六元内酰胺环断开,17c 号位上形成 硫醚 结构。
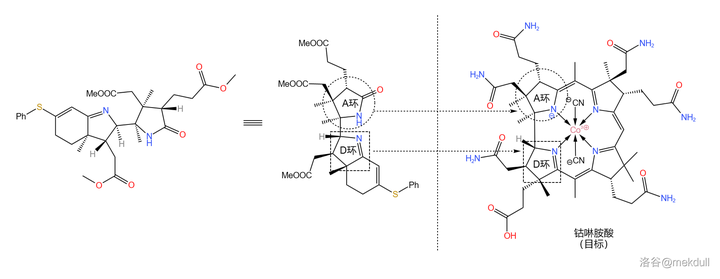
现在,我们手上的物质到底对应咕啉环的哪个部位已经可以看得很清楚了。如果你觉得不够清楚,那我们就把它旋转一下:
随后,在低温(-90^{\circ}C )的甲醇中用臭氧处理该物质,双键断裂,并在 17c 号位上生成 \ce{-COSR} 的结构。接下来,用液氨处理之,17c 号位上形成末端酰胺结构:
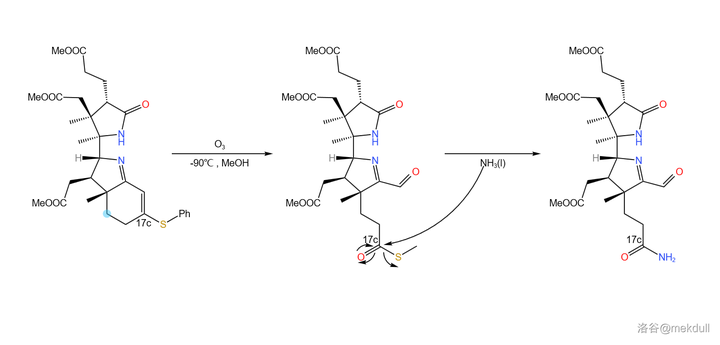
(注:其实 \ce{-COSH} 这个基团是有一个专有名的,读作you基,\ce{-COSR} 则名为you酯基,但我死活打不出来这个you字,还请各位指教;下文中我会把 \ce{-COSR} 称为「 硫酯 」,而把 \ce{-CSOR} 称为「 硫羰酯 」,请注意区分)
最后一步,在还原剂 硼氢化钠 (\ce{NaBp} )的作用下,15 号位上的醛被还原为醇;加入过量的 甲磺酸酐 (\ce{MsOMs} ),将醇甲磺酰化的同时为 17c 号位上的末端酰胺脱水,得到一个 氰基 (\ce{-CN} );最后加入 溴化锂 ,左半部分「cyanobromide 」的全合成就大功告成了:
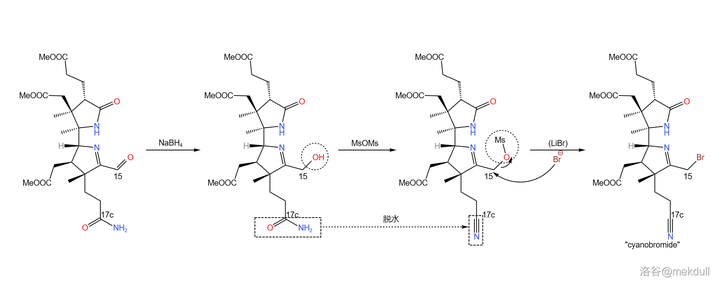
然而,这也只是一半而已。接下来,舞台交给 Eschenmoser 的课题组,他们将要带我们去看看,分子的右半部分(「thiodextrolin 」)是怎么合成出来的。
3.2 右半部分的合成
相比于 Woodward 课题组的工作,Eschenmoser 课题组设计的合成路线更为简短,但同样精妙。
右半部分的合成起始于三种结构简单、产量巨大的重要化工原料: 乙醛酸 (\ce{HOCCOOH} )、 丁酮 (\ce{CpCOCpCp} )和 丁二烯 (\ce{Cp=CHCH=Cp} )。前两者在酸催化下发生 交叉羟醛缩合反应 ,得到一种烯基酮酸;随后,在 四氯化锡 (\ce{SnCl4} ,作为 路易斯酸 )的催化下,该物质与丁二烯发生著名的 Diers-Alder 反应,得到一种环己烯的衍生物:
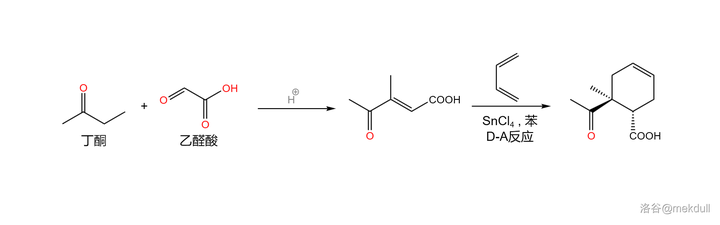
Diers-Alder 反应(又名 双烯加成反应 )在有机合成中非常重要,其机理在前几篇文章中已有提及,而在这篇文章的后记中,我还会简要说明全合成工作为这个反应带来的发展。
接下来,用过量的 Jones 试剂( 三氧化铬 (\ce{CrO3} )溶于 浓硫酸 )处理该物质,环状烯烃结构的双键随即断裂(此处反应条件非常强烈,一般来说 Jones 试剂不会氧化双键),生成一种带有三个羧基的中间体;该中间体随后 自发环化 ,形成 双内酯 结构,产率 75\% :
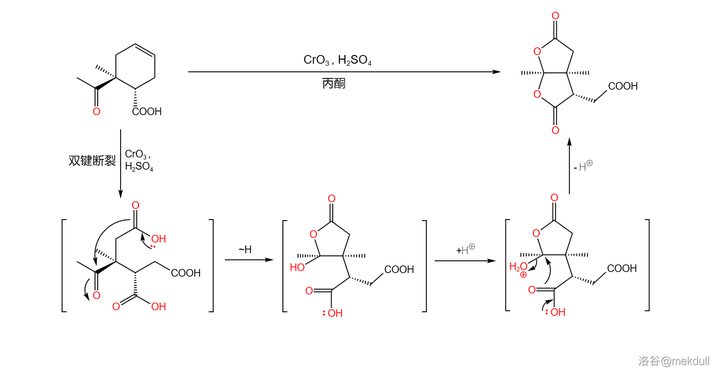
关于 \ce{Cr(VI)} 氧化剂断裂双键的机理,一般认为是先生成 环氧化合物 ,随后生成 邻二醇 ,最后在过量氧化剂的作用下,碳碳键断裂,得到二酸产物:
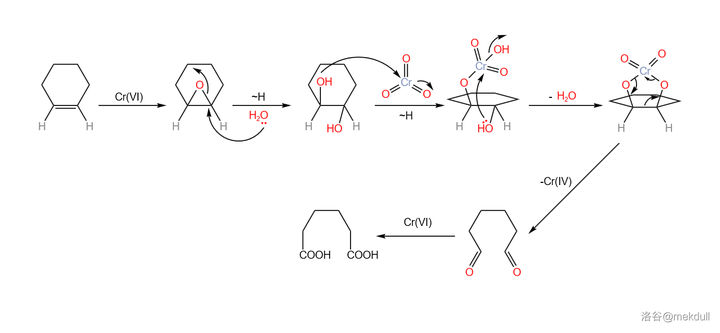
言归正传。下一步要用到的是 Arndt-Eistret 同系增碳反应,顾名思义,这个反应可以为羧基 增加一个同系碳原子 。在 \ce{SOCl2} 作用下,分子内的羧基首先变成酰氯;加入 \ce{CpN2} ,在亲核取代之后,同系碳原子随即被引入;在 \ce{Ag(I)} 的催化下,分子放出 \ce{N2} 形成 卡宾酮 中间体,随后发生重排反应,最终在 \ce{MeOH} 的作用下变成酯,三步反应总产率 69\% :
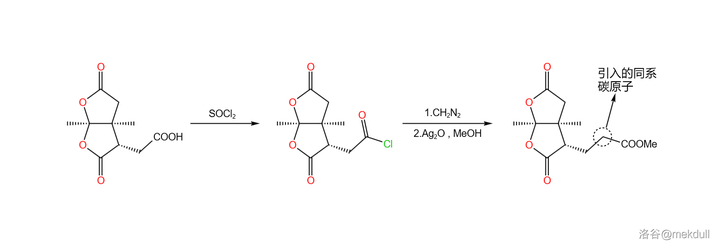
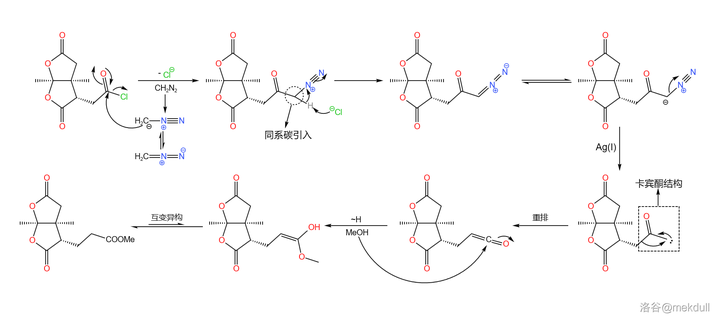
随后,在甲醇中用定量的 \ce{Np} 处理该物质,可以 将其中一个内酯基团转化为内酰胺 。由于特殊的 双内酯结构 ,该物质的羰基具有更强的亲电性,所以在室温下反应就可以快速进行:
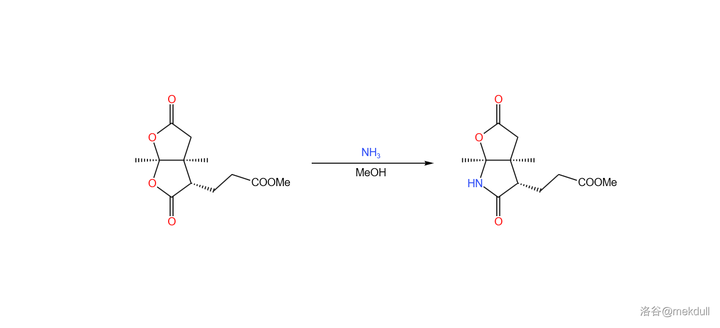
反应产率约为 55\% (其余的 45\% 大多都是另一个内酯基团被变成了内酰胺)。可见,这个反应的选择性其实并不显著。
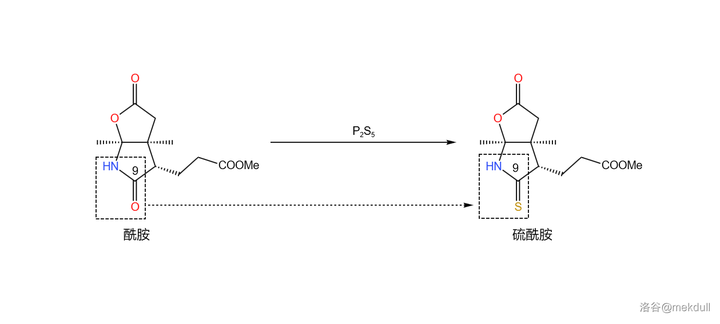
但不管怎样,55\% 的产率也算是能接受了。现在让我们 把产物分成两份 ,并对它们 进行不同的处理 。第一份的处理很简单,在 \ce{THF} 中,用一定量的 五硫化二磷 (\ce{P2S5} )进行硫化,9 号位的内酰胺结构被转化为 硫内酰胺 ,而分子中的其他酯基则不受影响:
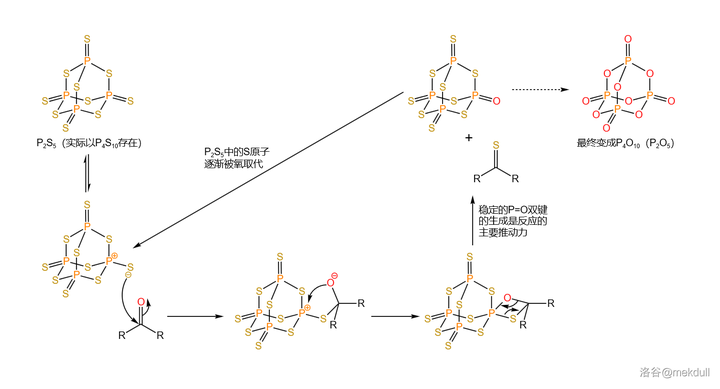
五硫化二磷 (\ce{P2S5} )的分子结构与所谓 五氧化二磷 (\ce{P2O5} )类似,实际上是以 \ce{P4S10} 形式存在的。作为一种重要的化工原料,它在有机合成中经常被用于将 羰基 转化为 硫羰基 。后来,人们又以它为基础制得了威力更强的版本,也就是著名的 劳森试剂 (Lawesson's~Reagent )。\ce{P2S5} 硫化的机理如下,有点类似于 Wittig 反应的机理:
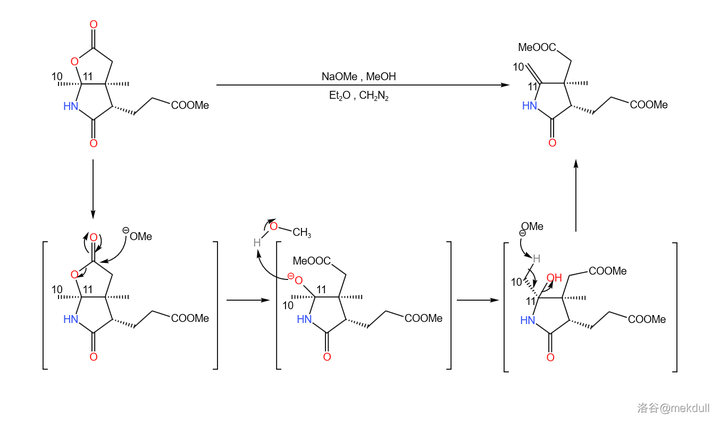
扯得有点远,让我们来看看第二份是怎么处理的。在 乙醚 (\ce{Et2O} )中,用 \ce{NaOMe} 裂解内酯环,随即消去 1 分子水生成 10,11 号位上的双键;为了防止需要的酯基被水解,反应体系中还添加有一定量的 \ce{CpN2} 。最终产率约为 \ce{92\%} :
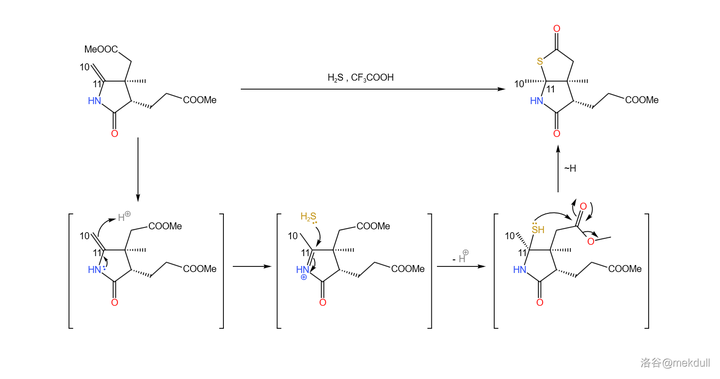
接下来,在 三氟乙酸 (\ce{CF3COOH} ,缩写 \ce{TFA} )的作用下,该物质与 \ce{pS} 加成(与加 \ce{pO} 的机理类似)。随后,硫原子进攻附近的酯基,形成五元环状硫酯(\ce{-COSR} )结构:
之所以要大费周章把它搞成硫酯,是因为在 威尔金森催化剂 (\ce{(PpP)3RhCl} )和 氰化钠 (\ce{NaCN} )的作用下,硫酯容易发生 脱硫化羰反应 ,从而把那个不需要的碳原子去掉;随后再用 \ce{Ag+} 处理,得到 10,11 号位上的双键。这个反应同时也会产生一种有三元环结构的副产物,因此产率并不是很高:
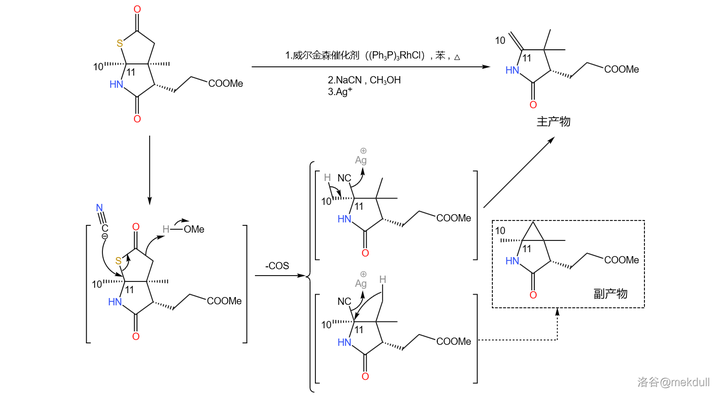
(注:由于涉及过渡金属催化,机理相当复杂,所以 此处展示的机理是简化过后的 。想要进一步了解的读者可以自行查阅相关文献。)
好,现在两份都已经完成了各自的处理,是时候把它们 结合起来 了。为了这一步,Eschenmoser 开创了一个全新的反应 ,也就是如今已被我们所熟知的 Eschenmoser 偶联反应。
Eschenmoser 偶联反应需要一个 硫酰胺 和一个 烯胺 。在 过氧化苯甲酰 (\ce{PhCOOOCOPh} )的作用下,两分子硫酰胺首先发生 氧化自偶联 ;随后,烯胺进攻自偶联产物, 二硫键 断裂得到 硫桥 结构,又自发异构化为 环硫化合物 ;最后在 \ce{P(III)} 化合物(此处使用 三乙氧磷 ,\ce{P(OEt)3} )作用下,硫原子被拔去,完成整个偶联反应:
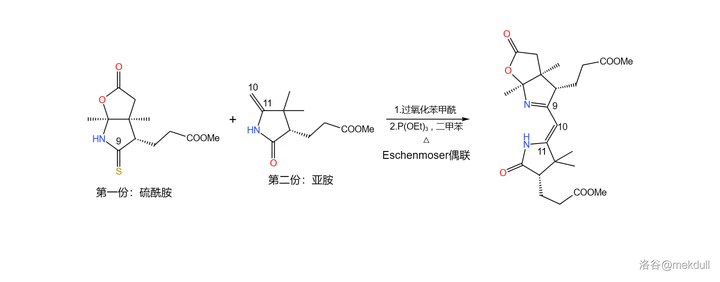
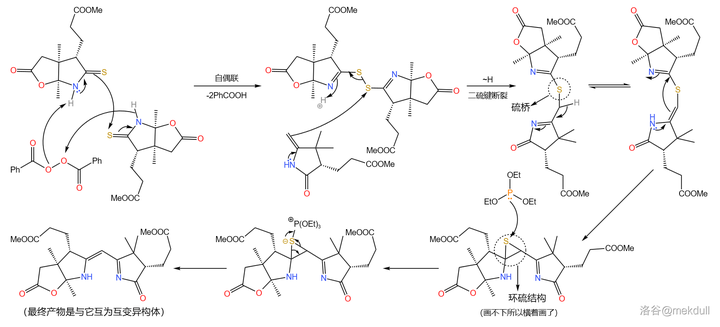
必须说,这个反应设计得非常巧妙,但它的 通用性并不好 ,因为底物之一的烯胺与亚胺是互变异构体,且一般 平衡偏向后者 。因此,有人对这个反应进行了改进,而稍后我们就会看到这种变异的 Eschenmoser 偶联反应。
偶联反应之后,距离完成 只差最后一步 了——将 14 号位的酰胺结构硫化。这一次,分子内多出了许多结构,为了保证它们不被破坏,课题组在多次实验后选择了在 甲基汞 离子(\ce{MeHg+} )催化下用 \ce{pS} 直接硫化的方案,并最终以高产率得到了目标产物:
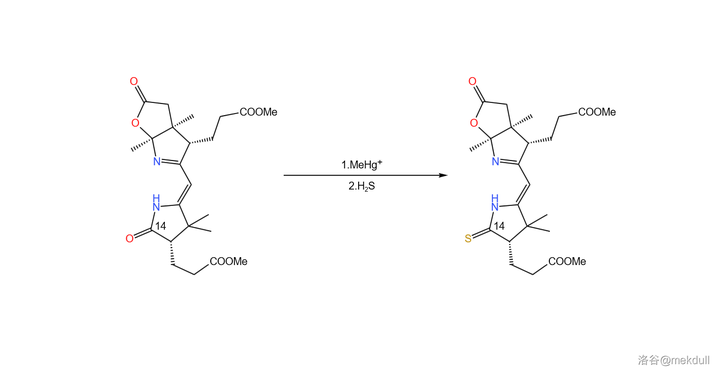
就这样,右半部分「thiodextrolin 」的全合成也宣告成功。接下来,就该把它们拼起来了。
3.3 咕啉环的拼接与修饰
这一部分的工作是 两个课题组各自完成 的,但篇幅所限,我们主要来讲一讲 Woodward 课题组设计的合成路线。(事实上,两条路线的 大体思路是基本一致的 )
拿到了左右两半后,首要的任务当然是将它们先拼在一起。Woodward 果断采用了 Eschenmoser 偶联的一种 变异反应 。这种变异将其中一种底物从烯胺变成了 活泼卤代烃 ,从而使反应的通用性大大增强;利用它,就能轻松地在 15 号位上完成偶联:
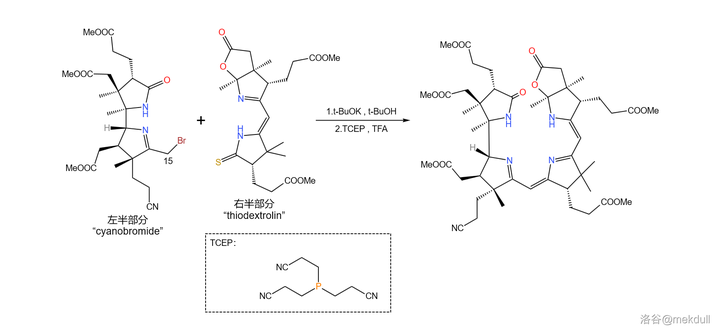
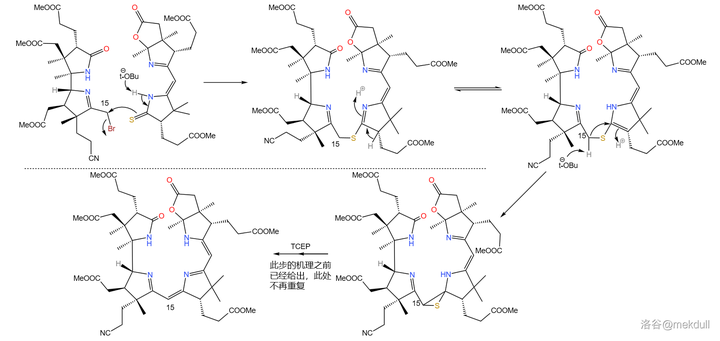
接着,在 4- 甲基吡啶 催化下用一定量的 \ce{P2S5} 处理之,4 号位的内酰胺和 7b 号位上活泼的内酯结构被硫化,分子中其他普通的酯基不受影响;随后加入 Meerwin 盐对 硫羰基 进行甲基化:
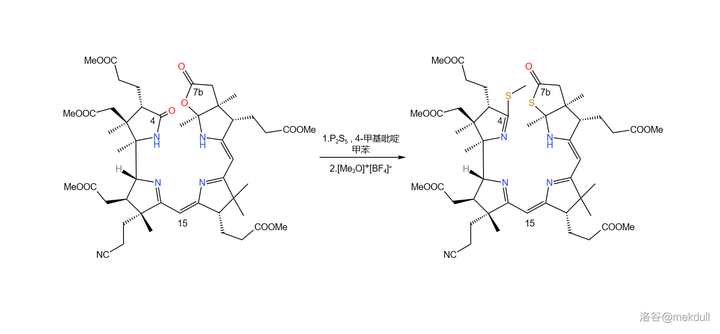
补充一句,\ce{P2S5} 在硫化酯基时一般先会生成 硫羰酯 ,也就是 \ce{-CSOR} 结构,但在碱性试剂作用下,硫羰酯会变成通常更稳定的 硫酯 (\ce{-COSR} )结构。对 7b 号位内酯结构的硫化就大致经历了这一历程。
下一步,使用 二甲胺 (\ce{Me2NH} )打开硫内酯环,得到 7b 号位的酰胺结构,同时在 5,6 号位上发生消去反应,生成双键。这个反应看似简单,但课题组却花费了不少时间去 修正反应条件 ,才使反应的产率足够令人满意:
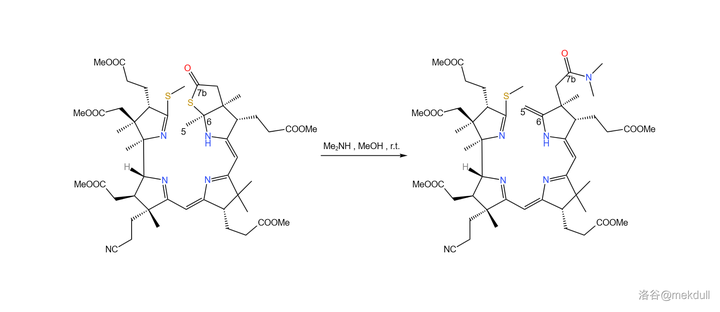
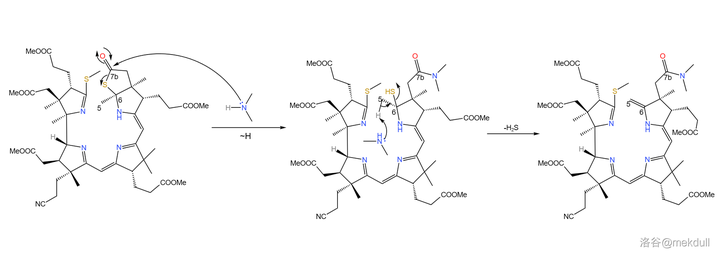
随后,课题组打算引入中心配位的 \ce{Co} 原子。一开始,课题组并没有预料到这一步的困难,但他们马上就会知道了:\ce{Co} 是一种 极其有效的催化剂 ,可以 迅速催化该物质分解 ,让前面的一切努力都变成白忙活。有人会问:为什么一定要这么着急地把 \ce{Co} 原子放进去呢?因为在下一步,我们将要进行 4 号位和 5 号位碳原子的偶联。还是由于 空间位阻 ,此时 4 号和 5 号碳原子虽然在纸面上看起来很近,但实际上距离很远——这就导致 分子内的偶联反应难以进行 。下图是由计算机渲染得到的,大家可以直观感受一下两者的距离:
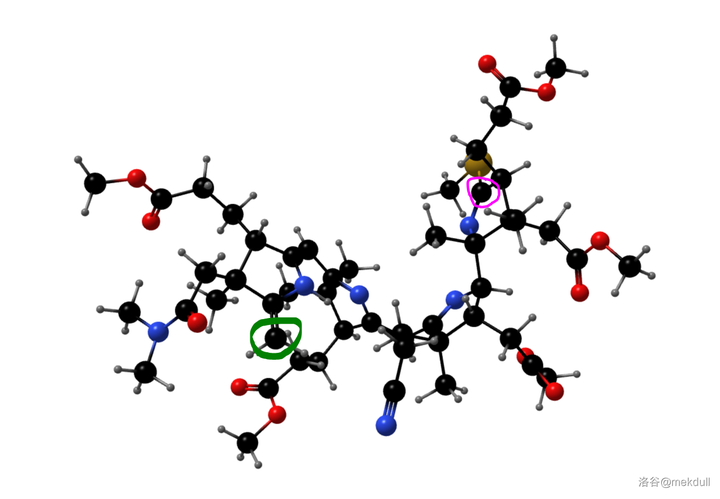
(上图:目前我们手中化合物的空间结构,绿色圈出的是 5 号碳原子,粉色圈出的是 4 号碳原子)
如果此时我们成功地将中心的 \ce{Co} 原子塞进去,那么 整个分子的空间结构都会因此改变 。4,5 两个碳原子会被强行「拉拢」,从而使得它们之间的偶联反应可以顺利进行。想到这里,Woodward 下定决心要攻克这道难题。它一定是有解的! 维生素B12就在那里 ,它的存在,就是问题有解的最好证明!带着这份信念,课题组花了 整整一年的时间 ,尝试各种方法和反应条件,同时也尝试揭秘 \ce{Co} 催化分解的机理。最终,他们找到了。
在最终的目标中,\ce{Co} 原子的氧化数应为 \ce{+III} ,但在几百次实验之后,课题组确信:正是 \ce{Co(III)} 破坏了分子的结构。所以,最终的解法就是:先在 \ce{THF} 中用 无水氯化钴 (\ce{CoCl2} )处理,让 \ce{Co(II)} 进入配位体系;随后加入 \ce{KCN} 水溶液,利用 \ce{CN-} 离子强大的配位能力把 \ce{Co(II)} 「锁」在分子的中心 ;等到万事俱备了,再将 \ce{Co(II)} 用空气氧化成 \ce{Co(III)} 。此时,就算是 \ce{Co(III)} 想搞破坏也不可能了:
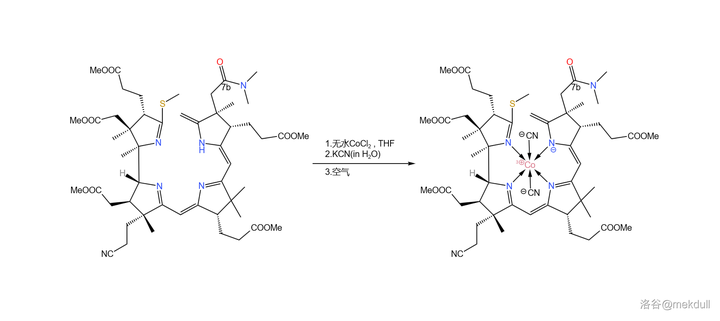
补充一句,此处的无水氯化钴也 不是随便选择的 。很早之前就有人发现,\ce{Cl-} 和 \ce{I-} 似乎对于 钴络合物 的形成具有催化作用,虽然这种作用的机理尚不明确。到了这里,如果不用无水卤化物的话,那么 \ce{Co(II)} 会很难进入配位体系,也就没法进行下一步的反应了。
难题解决之后,后面的反应就如同预料一般顺利了。在 \ce{DMAC} ( 二甲基乙酰胺 ,\ce{CpCON(Cp)2} )中用 有机碱 \ce{DBN} 进行催化,偶联反应就可以高产率地进行:
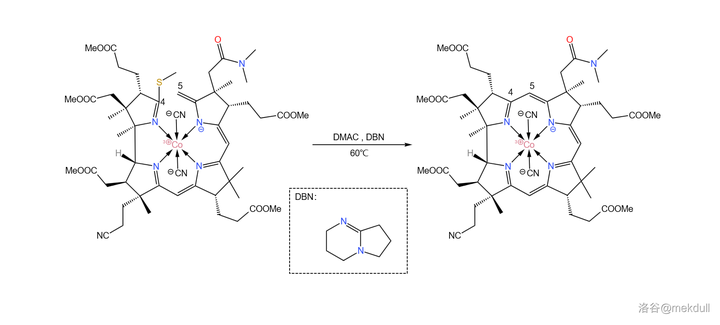
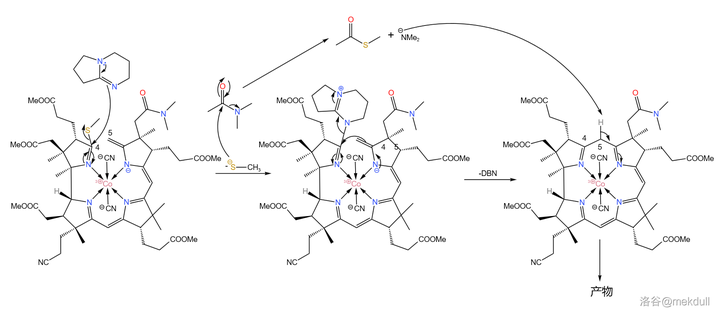
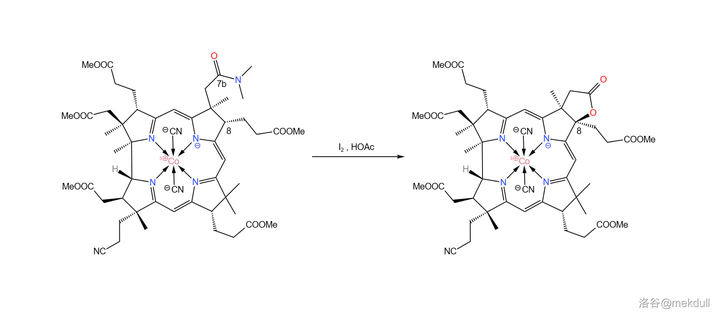
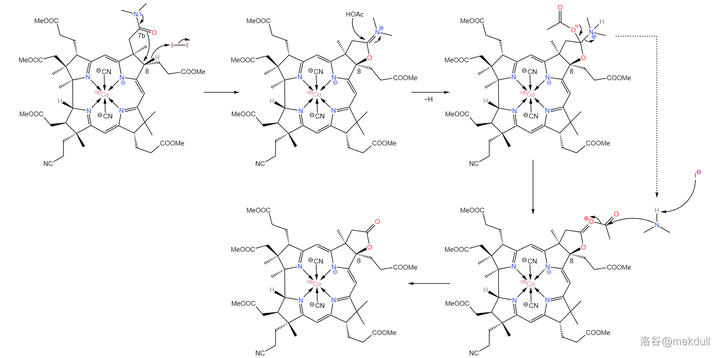
现在, 整个咕啉环的拼接已经宣告成功 ,接下来便是对其进行修饰。在 \ce{AcOH} 中用 碘单质 氧化该物质,8 号位的 \ce{C-H} 键断裂,分子右上角的 7b 号位的酰胺结构变成内酯:
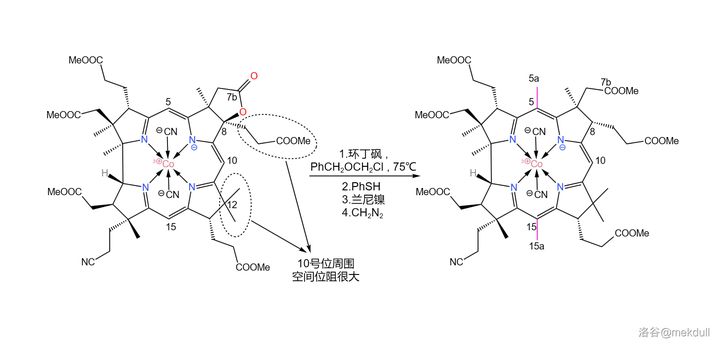
下一步,我们需要在 5 号位和 15 号位上各补上一个甲基(或许很多人都没有注意到这两个位置缺少一个甲基)。在 环丁砜 (溶剂)中,用 苄基氯甲基醚 (\ce{PhCpOCpCl} )可以直接活化 5,15 号位上的 \ce{C-H} 键并取代之;随后加入 苯硫醇 (\ce{PhSH} ),5a,15a 号碳原子上的 苄氧基 (\ce{-OCpPh} )随即被取代;接着,用 兰尼镍 (Raney~\ce{Ni} )进行 还原脱硫反应 ,7b 号位上的内酯结构也随即被破坏,游离出羧基;最后用 \ce{CpN2} 进行酯化,完成整个反应历程。
有人可能会问,为什么 10 号位上的氢原子没有被取代呢?观察分子的结构就能发现,10 号位周围的空间 非常拥挤 ,附近的 8,12 号位的碳原子都是 完全取代 的(其实,上一个反应的目的就是将 8 号位完全取代,从而增大空间位阻),就像是一道屏障一般阻隔了试剂对 10 号位的进攻。这足以保证这个反应的选择性:
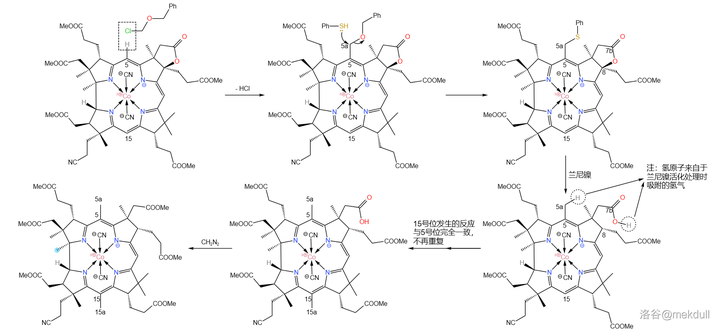
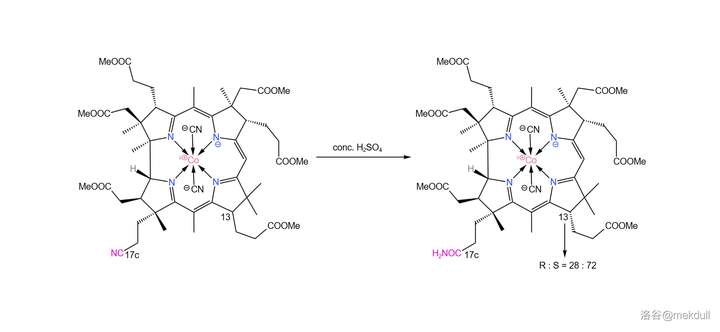
随后,我们需要把 17c 号位的氰基(\ce{-C#N} )水解成羧基(\ce{-COOH} )。由于分子中 有众多易水解的酯基 ,所以这并不是一项简单的任务。课题组选择先用浓硫酸处理该物质,将氰基 水解为末端酰胺 。没错,虽然一般浓硫酸是以「 脱水剂 」的身份出现的,但这里它扮演了一个相反的角色。这里的反应条件看似强烈,但主体的咕啉结构基本没有遭到破坏。
除此之外,浓硫酸还起到了一个意料之外的作用。分子中的 13 号碳原子可以通过 互变异构 的方式转变自己的构型,而浓硫酸恰好提供了这个条件。最终,13 号碳原子的 R:S 平衡为 28:72 (我们需要的是 S 型):
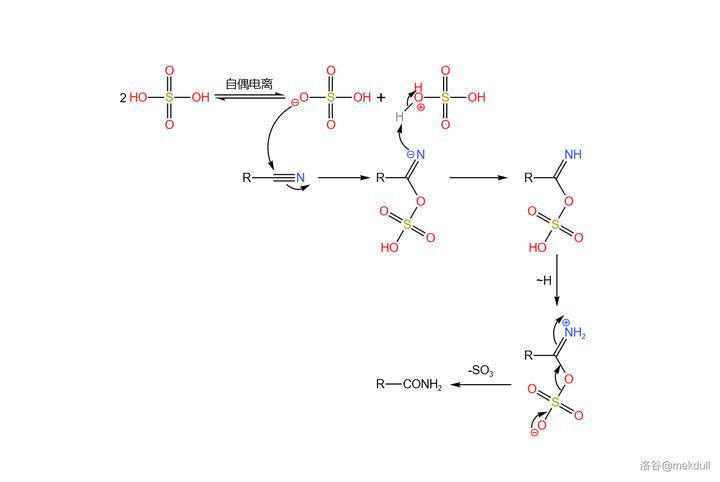
接下来就是考虑如何将酰胺结构水解成羧基。为了防止酯基被水解,课题组首先考虑了使用 亚硝酸酯 (通式 \ce{RONO} )作为水解试剂。遗憾的是,亚硝酸酯虽然出色地完成了水解任务,但它同时也会将分子的 10 号位 亚硝化 。为此,Woodward 课题组筛选了众多 亚硝酸衍生物 ,但都无济于事。最终,这一方案被放弃了。
在此关键时刻,隔壁的 Eschenmoser 课题组送来了助攻。简单来说,他们发明了一种强大的 亲电试剂 (在此就姑且称之为 Eschenmoser 亲电试剂吧),可以通过 氟硼酸银 (\ce{AgBF4} )与一种 环己胺衍生物 反应生成。在非常弱的酸性条件下(可以用 酸碱缓冲体系 ),它就可以将酰胺转化为 羟乙醛酯 。最后只需加入 \ce{Me2NH} ,就可以将活泼的羟乙醛酯转化为羧酸:
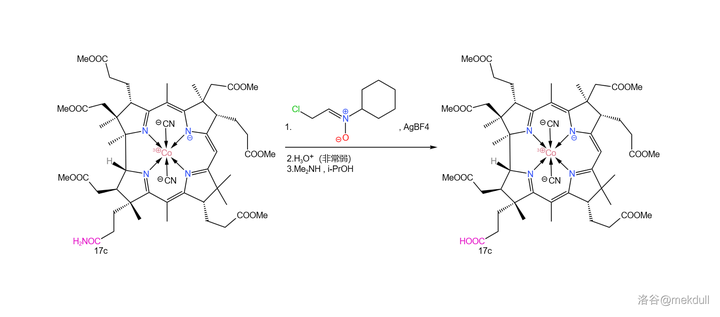
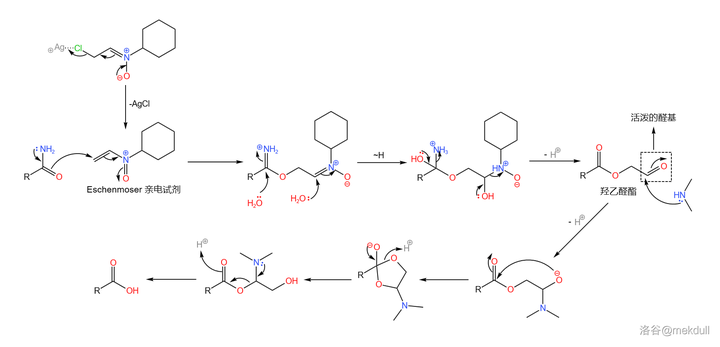
这个反应的设计极其巧妙,而且在面对如此复杂的底物时也可以给出超过 90\% 的产率——Woodward 评价这个反应「 狡猾而优雅 」。后续的研究证明,这个反应 对于几乎所有酰胺都是有效的 ,不仅限于末端酰胺。如今,我们一般把它称为 Eschenmoser 酰胺水解反应,在复杂天然产物的合成中常会用到。
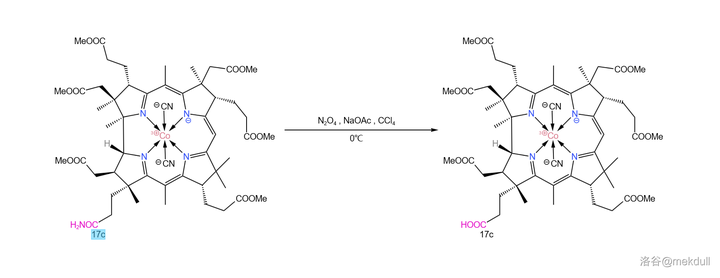
或许是不愿落后于人,Woodward 课题组很快也发现了这个问题的另一种解法:在 四氯化碳 (\ce{CCl4} )中用 \ce{NaOAc} 和 四氧化二氮 (\ce{N2O4} )代替亚硝酸酯,就可以将酰胺有效水解,同时分子的 10 号位不再被亚硝化。反应产率 \ce{70\%-80\%} ,虽然比 Eschenmoser 的方法差了点,但如果从经济上考虑,那 Woodward 的方法显然是更优的:
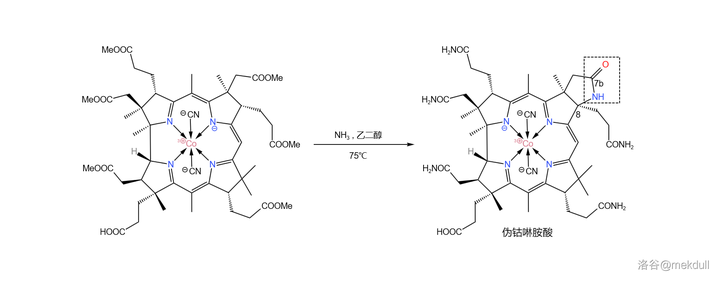
现在,距离成功合成钴啉胺酸只差一步了:将分子中的六个酯基全部转化为酰胺。在 75^{\circ}C 左右用氨(\ce{Np} )与 乙二醇 (\ce{HOCpCpOH} )大致 1:1 的混合物处理该物质,可以以高产率得到一种结晶。但令人大跌眼镜的是, 它根本不是钴啉胺酸 !
这种物质的 化学性质与钴啉胺酸几乎一致 ,且两者具有基本相同的 红外光谱 、 电子光谱 ,甚至于连 纸色谱 、 平面色谱 都完全无法区分二者。只有 高压液相色谱 系统告诉我们:它们不一样!如下图所示,课题组得到的物质在化学式上比钴啉胺酸少了两个 \ce{H} 。在它的右上角,7b 号碳原子形成了内酰胺结构。课题组将这种物质称为「 伪钴啉胺酸 」(pseudocobyric~acid ):
为什么分子会莫名其妙地去掉两个 \ce{H} ?难道是 有空气混进了反应体系 ?在反复检查之后,课题组排除了这种可能。那它到底为什么会被氧化呢?等等,分子中心还有一个配位的 \ce{Co} 原子!Woodward 猜想,中心配位的 \ce{Co(III)} 可以通过某种机制变成 \ce{Co(I)} 。由于 \ce{Co(I)} 具有很强的还原性,所以它会迫使分子会放出 \ce{p} ,并把自己变回 \ce{Co(III)} 。
所以,该怎么解决这个问题?一开始,课题组希望 找到一种合适的还原剂 ,将伪钴啉胺酸还原成钴啉胺酸。他们搜寻了好几个月,但根本没有收获。就在此时,他们意外地发现,只需将少量 铵盐 (比如 氯化铵 \ce{NpCl} )加入反应体系,效果就立竿见影。伪钴啉胺酸从产物中消失了,取而代之的是几乎 100\% 产率的钴啉胺酸:
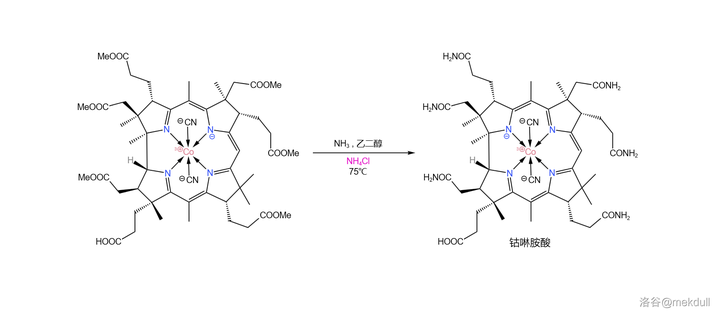
为什么铵盐会有这么神奇的效果?Woodward 认为,生成伪钴啉胺酸的机理中可能包含一个 酰胺负离子 中间体(\ce{-CONH^-} ),而 铵根离子 (\ce{Np+} )可以有效地 淬灭 酰胺负离子,从而抑制了伪钴啉胺酸的生成。
由于没有详细研究,Woodward 并没有在论文中给出生成伪钴啉胺酸的详细机理。但这或许已经不重要了,因为这一步的完成,标志着 维生素B12的全合成宣告成功 。这座「有机合成界的珠穆朗玛峰」,就这样在人们的齐心协力之下被征服了。这一年是 1972 年。
但是,我们的旅程还没有结束。毕竟,钴啉胺酸距离维生素B12 还差一个结构类似于核苷酸的侧链 ,那这个侧链又应该如何合成呢?
3.4 核苷酸侧链的合成与拼接
在概述中已经提到,这一部分的工作是德国化学家 Bernhauer~K. 等人在 1960 年完成的,所以对于 Woodward 来说,这部分的合成方法是现成的。但无论怎么说,这条侧链都是维生素B12的一部分,所以一篇完整的全合成介绍是少不了它的。
由于侧链的结构 类似于核苷酸 ,所以 Bernhauer 的课题组也采用了类似于核苷酸的合成策略。但值得注意的是,这条侧链的「 碱基 」并不是 \ce{A,C,G,T,U} 中的任何一个,而是一种 苯并咪唑 ; 磷酸基团 则是接在 核糖 结构的 3' 位,而不是 5' 位:
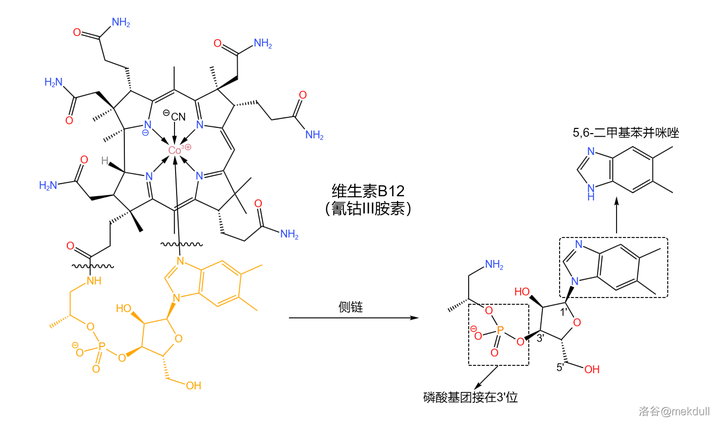
合成的起始物是一种有机染料工业的重要原料:4,5- 二甲基邻苯二胺 。在 多聚磷酸 (或其他较强的酸)的作用下,它可以与 甲酸 缩合得到 5,6- 二甲基苯并咪唑:
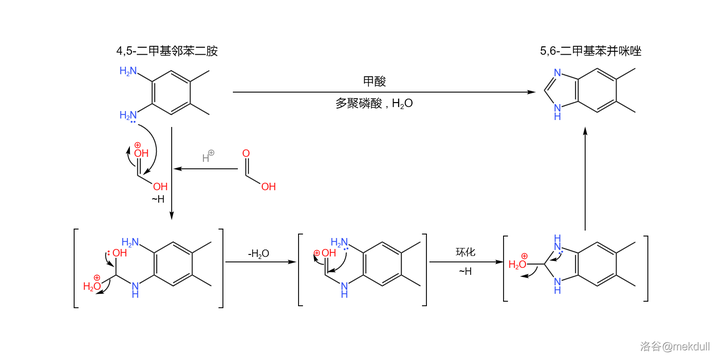
(注:较新的研究发现,上反应可以在 \ce{DMF} 中被 三甲基氯硅烷 (\ce{TMSCl} )催化,在此条件下产率可以达到 95\% 。上图中展示的反应条件是上世纪使用的,产率相对较低)
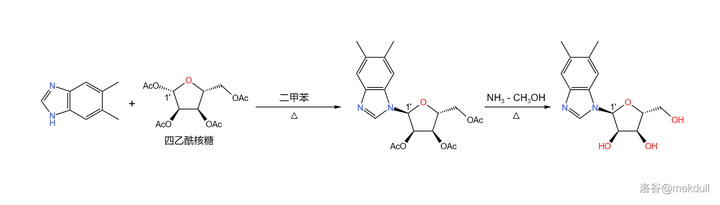
这样一来,碱基就搞定了。随后登场的是一种核糖的衍生物: 四乙酰核糖 ——它直到今天都是核苷酸合成中的重要试剂。在二甲苯中,它与刚才制得的 5,6- 二甲基苯并咪唑在 1' 号位发生 缩合反应 ;将反应的产物用 \ce{Np-CpOH} 处理,脱去多余的乙酰基后得到结构类似于 核苷 的物质:
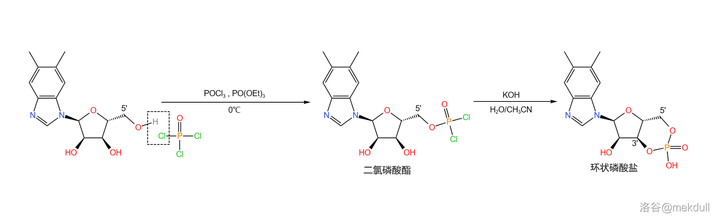
下一步,在低温下用 三氯氧磷 (\ce{POCl3} )和 磷酸三乙酯 (\ce{PO(OEt)3} ,做溶剂)处理该物质,得到 5' 号位的 二氯磷酸酯 结构;随后用强碱处理之,得到 3',5' 号位的 环状磷酸盐 :
接着,在很弱的碱性(\text{PH}\approx8.0 )条件下,用 铈离子 (\ce{Ce^3+} ,此处由于碱性太弱无法产生 氢氧化铈 (\ce{Ce(OH)3} )沉淀;最近有研究指出,实际上起作用的是 \ce{Ce^3+} 在此条件下部分水解产生的 簇状羟基配合物 )催化环状磷酸盐部分水解,可以得到 3' 号位的磷酸(副产 5' 位的磷酸):
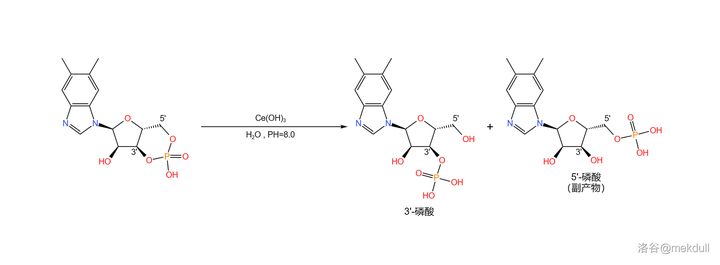
下一步,我们要把 3'- 磷酸与 异丙醇胺 (\ce{pNCpCHOHCp} ,有两种 旋光异构体 ,此处使用 R- 异丙醇胺)缩合成磷酸酯。为了防止氨基的干扰,课题组首先将其与 苯甲醛 (\ce{PhCHO} )反应,把氨基保护起来;随后,在强碱 正丁醇钠 (\ce{t-BuONa} )的作用下,两者顺利地发生缩合反应;反应结束后用乙酸水溶液脱去保护基,就能得到这条 完整的核苷酸侧链 :
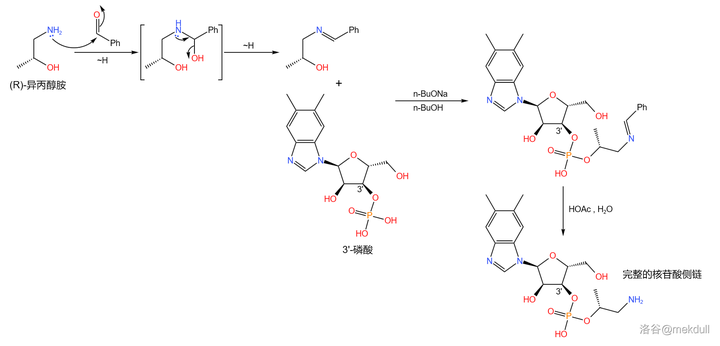
最后,我们需要 把这条侧链「装」到钴啉胺酸上 。为此,课题组测试了多种反应条件,最终选择使用 柠檬酸铵 溶液和 异戊醇 的混合物处理。二者顺利地发生脱水缩合反应,以 92\% 的产率得到最终产物:
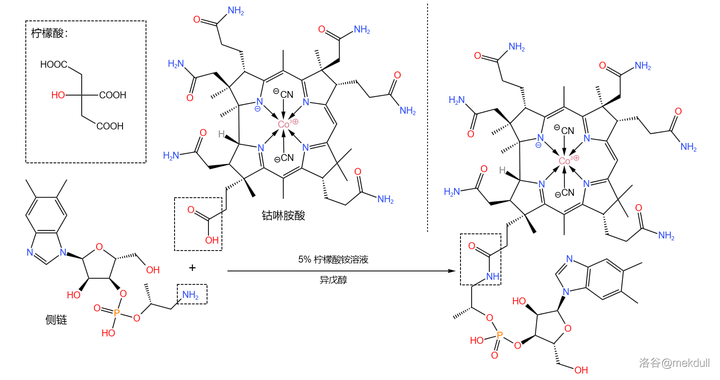
随后,该物质在此条件下可以自发消去一个 \ce{HCN} 分子,而侧链顶端的 \ce{N} 原子则会自动进入中心 \ce{Co} 原子的配位体系,从而得到成品维生素B12:
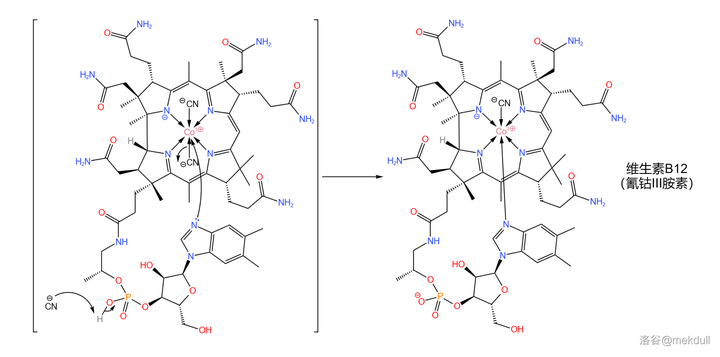
至此,那一抹亮丽的红色终于出现在了人们眼前。
4.总结与后记
这或许是全网最完整、详细的介绍维生素B12全合成的中文文章了。全文字数超过了一万五千,也有八十多张配图。如果喜欢还请多多支持,谢谢!(上次说的关于牛胰岛素全合成的解析已经在写了,但由于资料和时间问题,我也不敢说什么时候可以写完)
维生素B12的全合成是有机合成界前所未有的壮举 。毫不夸张的说,这项工作可以成为划分「 近现代 」有机化学和「 当代 」有机化学的分界线。它的意义远不止 拓展了有机合成复杂性的界限 ,更重要的是,它的成功沉重打击了几百年来「 化学局限论 」的思想,让人们开始相信: 化学真的可以为人类创造一个新的世界 。
即使抛开思想上的启迪,这项工作 在实际上的意义也不遑多让 。在此次全合成中首次亮相的 Eschenmoser 偶联反应已经成为 构筑碳碳键的重要手段 ,在许多药品的工业化合成中得到应用;Bernhauer~K. 等人合成侧链的方法在之后的岁月中被不断改进,衍生出了众多重要的 核苷酸化学合成法 ,为今日 腺苷 、 肌苷 、\ce{ADP} 、\ce{ATP} 等生物分子的大规模合成打下了基础,也成为了 遗传物质化学合成 的支点之一;咕啉环的人工合成法指导了人们探寻生物体内咕啉结构的构建,并最终将其 合理化 ,阐明了生物合成维生素B12的机理;Woodward 团队在合成过程中阐明了 互变异构导致的构型翻转 ,再一次提升了人类对于手性的认识,指导着后世几乎所有的手性合成方法;合成过程中发现的 \ce{Co} 元素的显著催化作用,到今天已经发展成为有机合成的一种强大策略,诸如 \ce{Co} 催化的 偶联反应、环加成反应、消去反应 等令人眼花缭乱;合成过程中,Woodward 和他的学生 Hoffmann 发现了 \ce{D-A} 反应在不同条件下的立体选择性,两人进而将这些规律总结、归纳,最终提出了名震整个有机化学界的 轨道对称守恒原理 ,也叫 Woodward-Hoffmann 规则——这被认为是近年来 化学理论方面的最大成就 。
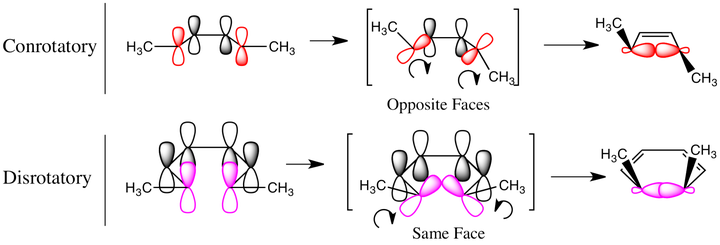
(上图:轨道对称守恒原理的简单图示)
在科学家们的努力之下,1993 年,维生素B12的 生物合成机制 被基本阐明。在此之后,人们开发出了维生素B12的 生物合成技术 ,这才有了今天能够直接在药店里买到的维生素B12药片。然而,谁能想到,在这种不起眼的廉价药品背后,暗藏着 150 余年来无数科学家为之不懈奋斗的史诗。
1979 年,积劳成疾的伍德沃德因心脏病突发去世,享年 62 岁。他在去世的前一天仍念念不忘那还未完成的 赤霉素 (gibberellin )全合成。最终,他的学生带领着剩下的课题组成员完成了这项工作;而在他去世 15 年后,他的另一位学生 Y.Kishi 将完成有机合成界「 新的珠穆朗玛峰 」—— 岩沙海葵毒素 (palytoxin )的全合成。如果伍德沃德能看到,想必他一定会倍感欣慰吧。
(超链接:关于岩沙海葵毒素全合成的详细解析)
相比于伍德沃德,另一位主角 Eschenmoser 教授并没有在这次伟大的工作中收获多少名声,如今或许也没有多少人记得他的团队也是维生素B12全合成的重要参与者。更鲜有人知的是,在完成这项工作之后,他选择 以维生素B12为切入点 ,研究两个根本看不到终点和希望的课题: 生命的化学起源 和 演化对于天然产物的构建 。尽管这注定是一条无比艰难的路,但他仍然义无反顾地走了下去。
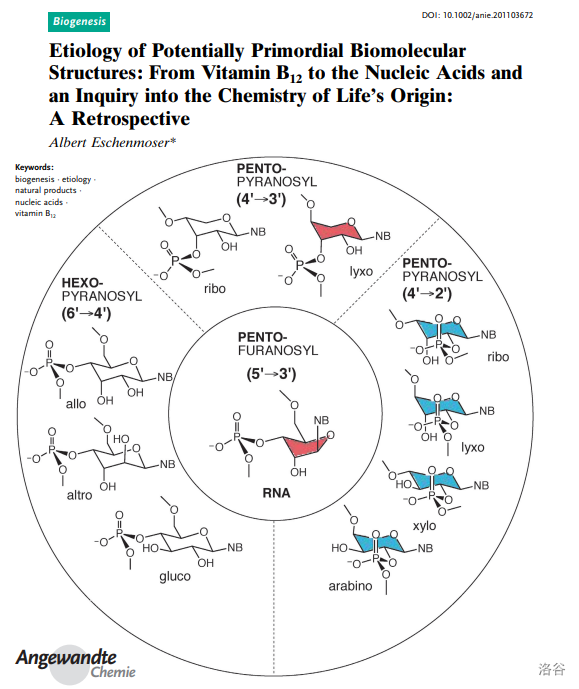
(上图:【潜在原始生物分子结构的病原学:从维生素B12到核酸,关于生命化学起源的探讨】论文影,A.~Eschenmoser ,2011 )
「我们可能永远也无从知晓」,这句老生常谈的话只会引导我们放弃寻找生命起源问题的努力······但对大自然创造生命之原理的探索永远不会真正结束,因为 这是我们在探索我们自己的起源,探索我们自己的曾经 。 ——【潜在原始生物分子结构的病原学:从维生素B12到核酸,关于生命化学起源的探讨】,A.~Eschenmoser ,2011A.~Eschenmoser 于 2023 年去世,享年 98 岁。显然,直到去世他也没有找到这两个终极问题的答案。但是,他的学生们接替了他,在这条看不到尽头的道路上继续前行。
伍德沃德曾经说过一句流传很广的话: 合成,是一门艺术 。但我认为,不光是有机合成, 任何科学探索,都是伟大的艺术 。
而艺术,需要人们不断地去传承与创新。
5.参考文献
[1] :Woodward R B. The total synthesis of vitamin B12[J]. Pure and Applied Chemistry Eighth, 1973, 33(1): 145-178.
[2] :Eschenmoser A, Wintner C E. Natural Product Synthesis and Vitamin B12: Total synthesis of vitamin B12 provided a framework for exploration in several areas of organic chemistry[J]. Science, 1977, 196(4297): 1410-1420.
[3] :Woodward R B. Recent advances in the chemistry of natural products[J]. Pure and Applied Chemistry, 1968, 17(3-4): 519-547.
[4] :Eschenmoser A. Studies on organic synthesis[C]//XXIIIrd International Congress of Pure and Applied Chemistry: special lectures presented at Boston, USA, 26-30 July 1971. Butterworths, 1971, 2: 69-106.
[5] :Friedrich W, Gross G, Bernhauer K, et al. Synthesen auf dem Vitamin‐B12‐Gebiet. 4. Mitteilung Partialsynthese von Vitamin B12[J]. Helvetica Chimica Acta, 1960, 43(3): 704-712.
[6] :Eschenmoser A. Vitamin B12: experiments concerning the origin of its molecular structure[J]. Angewandte Chemie International Edition in English, 1988, 27(1): 5-39.
[7] :Eschenmoser A. Etiology of potentially primordial biomolecular structures: from vitamin B12 to the nucleic acids and an inquiry into the chemistry of life’s origin: a retrospective[J]. Angewandte Chemie International Edition, 2011, 50(52): 12412-12472.
[8] :Friedrich W, Bernhauer K. Zur Konstitution des Vitamin B12-Faktors III[J]. Zeitschrift für Naturforschung B, 1954, 9(11): 685-697.
[9] :Woodward R B, Hoffmann R. Stereochemistry of electrocyclic reactions [J]. J Am Chem Soc, 1965, 87(2): 395-397.
[10] :Woodward R B, Hoffmann R. Conservation of orbital symmetry [J]. Acc Chem Res, 1968, 1(1): 17-22.
[11] :Zimmerman H E. A mechanistic analysis of the Birch reduction[J]. Accounts of chemical research, 2012, 45(2): 164-170.
[12] :KARUVALAM R, HARIDAS K, SHETTY S. Trimethylsilyl chloride catalyzed synthesis of substituted benzimidazoles using two phase system under microwave conditions, and their antimicrobial studies[J]. Journal of the Chilean Chemical Society, 2012, 57(2): 1122-1125.
[13] :Blanche F, Cameron B, Crouzet J, et al. Vitamin B12: How the problem of its biosynthesis was solved [J]. Angew Chem Int Ed, 1995, 34(4): 383-411.











