终于可以答高大上的进化生物学问题了,开心!
有人感觉文不对题,那先正面回答题主的问题吧。地球上最初的生命为什么有的进化成了植物有的进化成了动物?答案是不清楚,也许只是偶然的机缘巧合。知乎上充斥着大量的关于进化上的目的性的问题(扫一眼右边的相关问题就可见一斑)。为什么会产生XX物种?为什么会产生XX器官?首先,你们为什么认为一定有一个原因呢? 它可能就是某次突发事件的产物。 其次,现阶段还没有能力去研究这样的问题,所以进化生物学基本只关注 「怎么来的」「怎么运作」 ,如果事实逻辑链能够解释一些目的性的问题,可以进行解释。但意向性目的论上的 「为什么」, 那实在太玄了。
当然,欢迎大家对于这样的问题大开脑洞啊,只是它不属于科学的范畴。进 化上意向性目的论解释的难点在于,它假设了进化的背后有一个意向主体。 比如有人说植物的产生是单细胞动物为了直接获得光合作用的能力吞噬了蓝藻,内共生出了叶绿体。这句话后半句是非目的性的事实陈述,是现在科学可以尝试解决的问题,但针对前面半句,这个「为了」的意向主体的什么?单细胞动物怎么制定出了这一宏大的战略?意向性目的论的尽头是智慧设计论,是神创论,是上帝。
但是从题主的题目描述来看,题目描述里的问题是进化生物学讨论的范畴,因为它不涉及意向性的目的论。针对这一问题,我们首先需要定义什么是动物,什么是植物。传统意义上动植物包含了整个真核生物域,但实际上在现代系统分类研究视野中动物和植物的关系比想象的要远得多。所以我们先看看当代的动植物到底在整个真核生物中处于什么位置。其次,解决从哪一步出现动植物分离,需要从系统发育的角度找到动植物的最近共同祖先,并考察此过程之后的各种进化现象。
直到目前,由于真核生物大类群的系统发育关系还不清楚,动植物的最近共同祖先的位置还不能确定,但可以根据已有的证据推测它们分道扬镳可能要追溯到整个真核生物的共同祖先,缘起则是一次真核细胞吞噬蓝藻之后内共生形成叶绿体。没有吞噬的细胞中的一支进化成了动物,拥有叶绿体的细胞中的一支进化成了植物。
撕掉手边过时的教科书,刷新一下大家对整个真核生物大分类的三观吧,其实除了通常意义上的动物,植物和真菌,真核生物里面还隐藏着数量远超过你想象的类群。有些种类稀少,不引人注意的类群,就像从天而降的外星生物,我们到现在都不知道它们是从哪里来的。
欢迎来到进化生物学和大系统发育研究的最前沿,这里面水很深,到现在都有很多一片漆黑的谜团无法得到澄清。先上一张这个领域的最新的成果,来自今年,采用了目前我们系统发育研究所能采用的最强大的工具——基因组(Katz & Grant, 2015)。整个系统发生树包含了150个基因,232种真核生物,36346个位点。
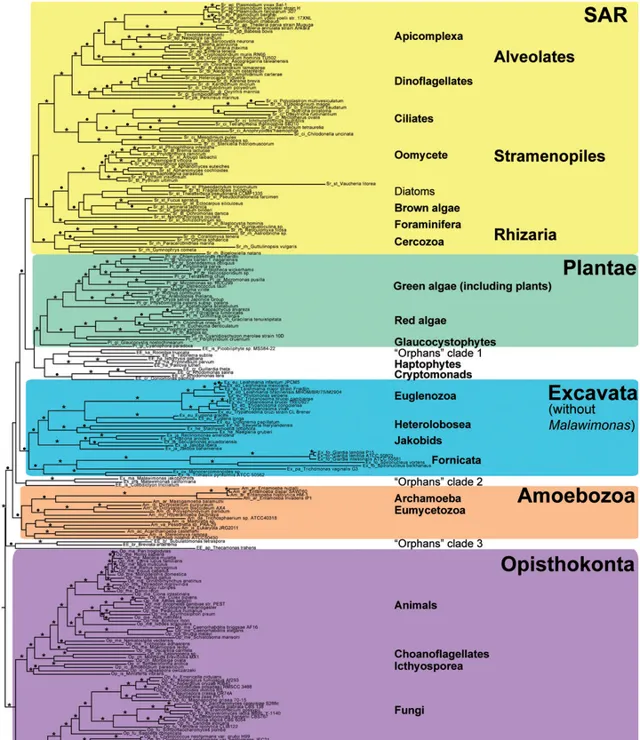
看不懂不要紧,我依次来详细说说。树上面五个不同颜色的大块标记了该项研究所发现的五个支持率高的单系类群(也就是可以被承认为符合真实进化历史的类群),分别为, SAR超类群、植物(Plantae)、古虫(Excata)、变形虫(Amoebozoa)、后鞭毛生物(Opisthokonta,包括真菌和动物) 。而那三个没有染色的空隙,就是三个种系发生不明的孤儿分支,我们还不知道它们是从哪里来的。从上到下依次说说这些都是什么生物。
SAR超类群 :名字来自这个类群里面三个大类群的首字母,分别是 囊泡虫类(Alveolates)、 不等鞭毛类(Stramenopiles)和 有孔虫( Rhizaria) :
囊泡虫类(Alveolates) :有自养也有异养生物,共同特征是存在皮层内生膜囊(cortical alveoli), 扁平的囊泡折叠为连续层支撑着膜,通常形成灵活的薄皮。成员来自以前的一部分广义植物和一部分原生生物。代表种为甲藻和疟原虫

叉状甲藻 Ceratium furca
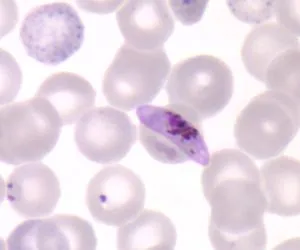
恶性疟原虫 Plasmodium falciparum
不等鞭毛类(Stramenopiles): 有自养也有异养生物。其生命周期的能动阶段,鞭毛虫细胞具有两种不同形状的鞭毛。成员分别来自以前的广义植物和真菌,代表物种为除绿藻红藻以外的大部分藻类和水霉。

海带 Laminaria japonica
某种水霉 Saprolegnia mixta
有孔虫( Rhizaria) :来自一部分以前的海洋原生生物。可分泌碳酸钙质外壳,但原生质为不定形变形虫态。有些种类可以保留吃掉的藻类的叶绿体进行光合作用(Bernhard,1999),为叶绿体二次内共生假说的可能性提供了某些参考。代表种为各类有孔虫。

Ammonia tepida

各种有孔虫骨骼。
SAR超类群中三个类群的单系支持率都很高,可以作为被广泛接受的类群,但三个类群之间的系统发生关系还不清楚。
泛植物(Plantae) : 大部分自养,包括各类陆生植物、绿藻、红藻和灰胞藻(Glaucophyte)。泛植物的叶绿体只包有两层膜,可能来自于直接吞噬蓝藻形成的内共生。而其他类群的叶绿体大多有三到四层膜,可能来自于吞噬绿藻和红藻的二次内共生。很有意思的是灰胞藻的叶绿体有肽聚糖层,可能是吞噬蓝藻形成内共生的遗迹(Keeling,2004)。
喜闻乐见的植物图片就不放了,放一张灰胞藻。
Glaucocystis sp.
古虫(Excavate): 包括自养或异养的一部分以前的原生生物,有很多寄生类原虫。共同特征是腹侧进食沟和联合细胞骨架结构。代表生物为可进行光合作用的眼虫和锥虫、滴虫。滴虫类不同寻常地缺少了典型线粒体,营寄生生活。

某种眼虫 Euglena sp.
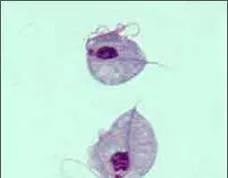
滴虫 Trichomonas sp.
变形虫类(Amoebozoa): 一类不定性原生质体。包括变形虫和黏菌。

某变形虫 Chaos carolinensis

煤绒菌 Fuligo septica
后鞭毛生物(Opisthokonta) :包括两类支持率良好的单系群: 菌物总界(Holomycota)和动物总界(Holozoa) 。共同特征为鞭毛细胞都具有单一个位于后侧的鞭毛。比如人类的精子。在大部分真菌里面鞭毛细胞已经消失,但在最低等真菌 壶菌(Chytrid) 中游动孢子仍然有后鞭毛。
动物总界(Holozoa): 值得关注的是狭义动物的几个姐妹群,来源于以前的一些原生生物和真菌。 领鞭毛虫( Choanoflagellate) :动物最近的亲戚。单细胞水生原生生物,触手呈领状环绕鞭毛,与最原始的动物之一海绵的领细胞基本结构相同,可以佐证它跟动物的近缘关系。

某领鞭毛虫 Monosiga brevicollis
DRIP小分支: 名字来自该分支的四个类群首字母。一类具有真菌特征的寄生微生物,动物的远方亲戚,种类包括肤胞虫(Dermocystida),鱼孢霉(Ichthyophonida)等。
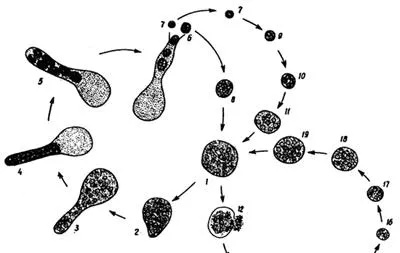
鱼孢霉 Ichthyophonus hoferi 的生活史。是一类重要的鱼类病害的病原体。
最后来看看那些空隙里的神秘类群:泛植物旁边的孤儿分支1里面包括一些小型光合生物。古虫旁边的孤儿分支2包括以前属于古虫的 马拉维单胞虫( Malawimonas ) .另一篇关于古虫界分类的文章(Hampl et al., 2009)也支持了马拉维单胞虫跟其它古虫的并系关系。变形虫旁边的孤儿分支3则包含了一系列单细胞的异养生物。这些地位未定的孤儿分支可能是进化中的孑遗分支,值得进行进一步的研究。而这些空隙的存在,也说明我们还将有可能发现新的生物类群。
大类群本身的单系性支持良好,但大类群之间的关系并不明朗。唯一可以作定论,且有其它研究作佐证的结论是,广义动物和广义真菌是单系群,且互为姐妹群。也就是说动物最近的亲戚是真菌。
关于大类群之间的关系有很多假说,我最喜欢的这一款:
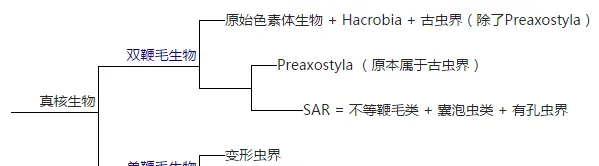
喜欢的原因大抵只是一个搞系统生物学的对于这样纯粹的二歧树的喜爱吧,简单的通过鞭毛细胞的鞭毛数量把大类群分为两类。并系和复系必须死!
双鞭毛那一支一直处于极度混乱的状态,暗无天日。单鞭毛生物一支情况要明朗一些,变形虫界和后鞭毛生物都是支持率较高的单系群,真菌和动物关系最为明朗,也有分子研究(Hampl et al., 2009)对单鞭毛生物这一支给出了较高的支持率。然而包括最新的那篇研究(Katz & Grant, 2015)和其它一些研究(Parfery et al., 2010)却显示出变形虫界可能跟其它双鞭毛生物关系更近,导致这一假说的命运又变得不明朗起来。
生物进化树的完美构建,这是从达尔文开始就萦绕在每个进化生物学家心中的终极梦想。当天体物理学家仰望浩瀚的星空的时候,进化生物学家则环顾着周围的草木鸟兽,信誓旦旦地说:我一定要让你们回到原本的位置上去!
然后理想是美好的,现实是艰难的,生物大系统树的构建到现在仍然是一个棘手的难题。即便应用了最先进的方法,仍然得不到令人满意的答案,期待以后能够有方法学上的突破。

来一个最新一期MBE的封面,每个系统进化生物学家的梦想
嘛有空可以补充一点空隙里面那些神秘类群的资料。











