終於可以答高大上的前進演化生物學問題了,開心!
有人感覺文不對題,那先正面回答題主的問題吧。地球上最初的生命為什麽有的前進演化成了植物有的前進演化成了動物?答案是不清楚,也許只是偶然的機緣巧合。知乎上充斥著大量的關於前進演化上的目的性的問題(掃一眼右邊的相關問題就可見一斑)。為什麽會產生XX物種?為什麽會產生XX器官?首先,你們為什麽認為一定有一個原因呢? 它可能就是某次突發事件的產物。 其次,現階段還沒有能力去研究這樣的問題,所以前進演化生物學基本只關註 「怎麽來的」「怎麽運作」 ,如果事實邏輯鏈能夠解釋一些目的性的問題,可以進行解釋。但意向性目的論上的 「為什麽」, 那實在太玄了。
當然,歡迎大家對於這樣的問題大開腦洞啊,只是它不屬於科學的範疇。進 化上意向性目的論解釋的難點在於,它假設了前進演化的背後有一個意向主體。 比如有人說植物的產生是單細胞動物為了直接獲得光合作用的能力吞噬了藍藻,內共生出了葉綠體。這句話後半句是非目的性的事實陳述,是現在科學可以嘗試解決的問題,但針對前面半句,這個「為了」的意向主體的什麽?單細胞動物怎麽制定出了這一宏大的戰略?意向性目的論的盡頭是智慧設計論,是神創論,是上帝。
但是從題主的題目描述來看,題目描述裏的問題是前進演化生物學討論的範疇,因為它不涉及意向性的目的論。針對這一問題,我們首先需要定義什麽是動物,什麽是植物。傳統意義上動植物包含了整個真核生物域,但實際上在現代系統分類研究視野中動物和植物的關系比想象的要遠得多。所以我們先看看當代的動植物到底在整個真核生物中處於什麽位置。其次,解決從哪一步出現動植物分離,需要從系統發育的角度找到動植物的最近共同祖先,並考察此過程之後的各種前進演化現象。
直到目前,由於真核生物大類群的系統發育關系還不清楚,動植物的最近共同祖先的位置還不能確定,但可以根據已有的證據推測它們分道揚鑣可能要追溯到整個真核生物的共同祖先,緣起則是一次真核細胞吞噬藍藻之後內共生形成葉綠體。沒有吞噬的細胞中的一支前進演化成了動物,擁有葉綠體的細胞中的一支前進演化成了植物。
撕掉手邊過時的教科書,重新整理一下大家對整個真核生物大分類的三觀吧,其實除了通常意義上的動物,植物和真菌,真核生物裏面還隱藏著數量遠超過你想象的類群。有些種類稀少,不引人註意的類群,就像從天而降的外星生物,我們到現在都不知道它們是從哪裏來的。
歡迎來到前進演化生物學和大系統發育研究的最前沿,這裏面水很深,到現在都有很多一片漆黑的謎團無法得到澄清。先上一張這個領域的最新的成果,來自今年,采用了目前我們系統發育研究所能采用的最強大的工具——基因組(Katz & Grant, 2015)。整個系統發生樹包含了150個基因,232種真核生物,36346個位點。
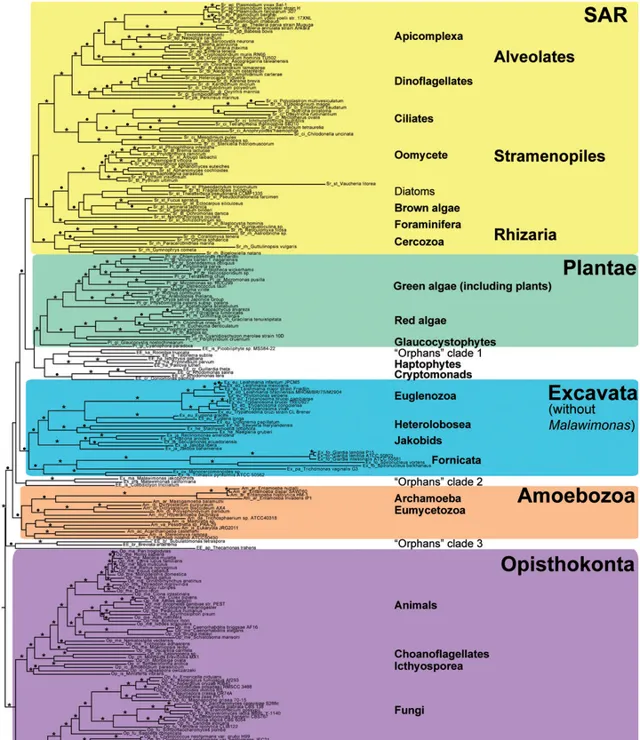
看不懂不要緊,我依次來詳細說說。樹上面五個不同顏色的大塊標記了該項研究所發現的五個支持率高的單系類群(也就是可以被承認為符合真實前進演化歷史的類群),分別為, SAR超類群、植物(Plantae)、古蟲(Excata)、變形蟲(Amoebozoa)、後鞭毛生物(Opisthokonta,包括真菌和動物) 。而那三個沒有染色的空隙,就是三個種系發生不明的孤兒分支,我們還不知道它們是從哪裏來的。從上到下依次說說這些都是什麽生物。
SAR超類群 :名字來自這個類群裏面三個大類群的首字母,分別是 囊泡蟲類(Alveolates)、 不等鞭毛類(Stramenopiles)和 有孔蟲( Rhizaria) :
囊泡蟲類(Alveolates) :有自營也有異營生物,共同特征是存在皮層內生膜囊(cortical alveoli), 扁平的囊泡折疊為連續層支撐著膜,通常形成靈活的薄皮。成員來自以前的一部份廣義植物和一部份原生生物。代表種為甲藻和瘧原蟲

叉狀甲藻 Ceratium furca
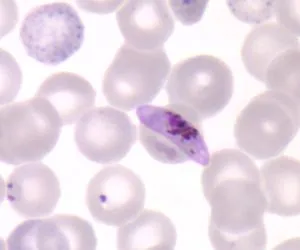
惡性瘧原蟲 Plasmodium falciparum
不等鞭毛類(Stramenopiles): 有自營也有異營生物。其生命周期的能動階段,鞭毛蟲細胞具有兩種不同形狀的鞭毛。成員分別來自以前的廣義植物和真菌,代表物種為除綠藻紅藻以外的大部份藻類和水黴。

海帶 Laminaria japonica
某種水黴 Saprolegnia mixta
有孔蟲( Rhizaria) :來自一部份以前的海洋原生生物。可分泌碳酸鈣質外殼,但原生質為不定形變形蟲態。有些種類可以保留吃掉的藻類的葉綠體進行光合作用(Bernhard,1999),為葉綠體二次內共生假說的可能性提供了某些參考。代表種為各類有孔蟲。

Ammonia tepida

各種有孔蟲骨骼。
SAR超類群中三個類群的單系支持率都很高,可以作為被廣泛接受的類群,但三個類群之間的系統發生關系還不清楚。
泛植物(Plantae) : 大部份自營,包括各類陸生植物、綠藻、紅藻和灰胞藻(Glaucophyte)。泛植物的葉綠體只包有兩層膜,可能來自於直接吞噬藍藻形成的內共生。而其他類群的葉綠體大多有三到四層膜,可能來自於吞噬綠藻和紅藻的二次內共生。很有意思的是灰胞藻的葉綠體有肽聚糖層,可能是吞噬藍藻形成內共生的遺跡(Keeling,2004)。
喜聞樂見的植物圖片就不放了,放一張灰胞藻。
Glaucocystis sp.
古蟲(Excavate): 包括自營或異營的一部份以前的原生生物,有很多寄生類原蟲。共同特征是腹側進食溝和聯合細胞骨架結構。代表生物為可進行光合作用的眼蟲和錐蟲、滴蟲。滴蟲類不同尋常地缺少了典型線粒體,營寄生生活。

某種眼蟲 Euglena sp.
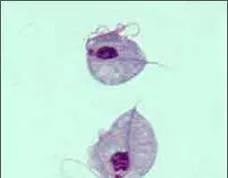
滴蟲 Trichomonas sp.
變形蟲類(Amoebozoa): 一類不定性原生質體。包括變形蟲和黏菌。

某變形蟲 Chaos carolinensis

煤絨菌 Fuligo septica
後鞭毛生物(Opisthokonta) :包括兩類支持率良好的單系群: 菌物總界(Holomycota)和動物總界(Holozoa) 。共同特征為鞭毛細胞都具有單一個位於後側的鞭毛。比如人類的精子。在大部份真菌裏面鞭毛細胞已經消失,但在最低等真菌 壺菌(Chytrid) 中遊動孢子仍然有後鞭毛。
動物總界(Holozoa): 值得關註的是狹義動物的幾個姐妹群,來源於以前的一些原生生物和真菌。 領鞭毛蟲( Choanoflagellate) :動物最近的親戚。單細胞水生原生生物,觸手呈領狀環繞鞭毛,與最原始的動物之一海綿的領細胞基本結構相同,可以佐證它跟動物的近緣關系。

某領鞭毛蟲 Monosiga brevicollis
DRIP小分支: 名字來自該分支的四個類群首字母。一類具有真菌特征的寄生微生物,動物的遠方親戚,種類包括膚胞蟲(Dermocystida),魚孢黴(Ichthyophonida)等。
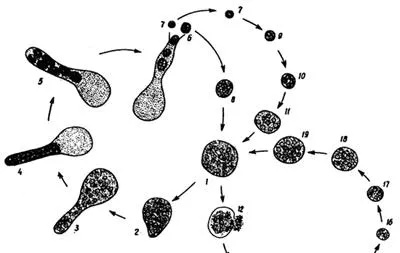
魚孢黴 Ichthyophonus hoferi 的生活史。是一類重要的魚類病害的病原體。
最後來看看那些空隙裏的神秘類群:泛植物旁邊的孤兒分支1裏麪包括一些小型光合生物。古蟲旁邊的孤兒分支2包括以前屬於古蟲的 馬拉維單胞蟲( Malawimonas ) .另一篇關於古蟲界分類的文章(Hampl et al., 2009)也支持了馬拉維單胞蟲跟其它古蟲的並系關系。變形蟲旁邊的孤兒分支3則包含了一系列單細胞的異營生物。這些地位未定的孤兒分支可能是前進演化中的孑遺分支,值得進行進一步的研究。而這些空隙的存在,也說明我們還將有可能發現新的生物類群。
大類群本身的單系性支持良好,但大類群之間的關系並不明朗。唯一可以作定論,且有其它研究作佐證的結論是,廣義動物和廣義真菌是單系群,且互為姐妹群。也就是說動物最近的親戚是真菌。
關於大類群之間的關系有很多假說,我最喜歡的這一款:
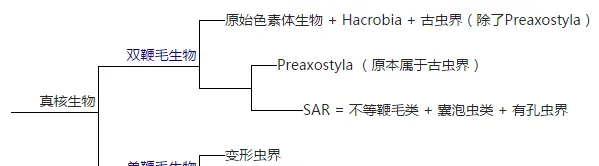
喜歡的原因大抵只是一個搞系統生物學的對於這樣純粹的二歧樹的喜愛吧,簡單的透過鞭毛細胞的鞭毛數量把大類群分為兩類。並系和復系必須死!
雙鞭毛那一支一直處於極度混亂的狀態,暗無天日。單鞭毛生物一支情況要明朗一些,變形蟲界和後鞭毛生物都是支持率較高的單系群,真菌和動物關系最為明朗,也有分子研究(Hampl et al., 2009)對單鞭毛生物這一支給出了較高的支持率。然而包括最新的那篇研究(Katz & Grant, 2015)和其它一些研究(Parfery et al., 2010)卻顯示出變形蟲界可能跟其它雙鞭毛生物關系更近,導致這一假說的命運又變得不明朗起來。
生物前進演化樹的完美構建,這是從達爾文開始就縈繞在每個前進演化生物學家心中的終極夢想。當天體物理學家仰望浩瀚的星空的時候,前進演化生物學家則環顧著周圍的草木鳥獸,信誓旦旦地說:我一定要讓你們回到原本的位置上去!
然後理想是美好的,現實是艱難的,生物大系統樹的構建到現在仍然是一個棘手的難題。即便套用了最先進的方法,仍然得不到令人滿意的答案,期待以後能夠有方法學上的突破。

來一個最新一期MBE的封面,每個系統前進演化生物學家的夢想
嘛有空可以補充一點空隙裏面那些神秘類群的資料。











