正式因為我們祖先守住了五根手指,我們才逆襲成為靈長之長。
當你無意中看到自己的五根手指,你可能想不到,決定你五根手指的基因來自3.5億年前的古老石炭紀。
如果你的手指是一個單獨的物種,它就是典型的活化石。
4.19億年前,夢幻鬼魚 [1] (Guiyu oneiros)在水裏自由自在遊弋時,恐怕不會想到,它們其中一支後裔會發展出四肢和指/趾骨,征服它們從來沒有踏足過的大陸,創造一段又一段演化傳奇。
夢幻鬼魚是最早的硬骨魚之一,同時也處在肉鰭魚演化樹的基幹位置(最底部)。
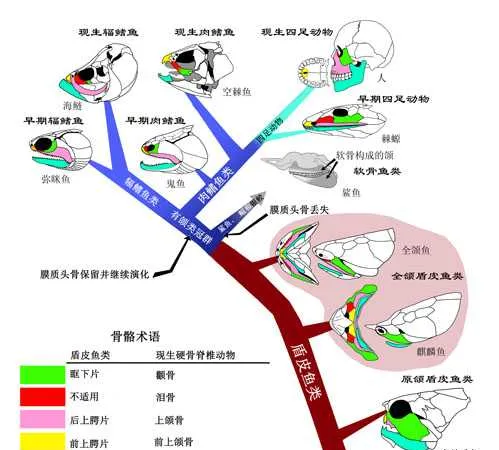
我們的上肢和下肢正是來源於它們的胸鰭和腹鰭。
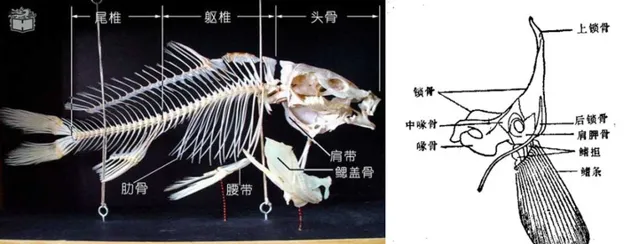
可以看出,魚鰭的主要結構便是鰭條。
很多人可能會理所當然地認為:
人類的手指,應該是由魚的鰭條演化而來,或者是一種全新發展出來的結構。
其實早期生物學家也這麽想,但卻被後來的研究打了臉 [2] [3] [4] 。
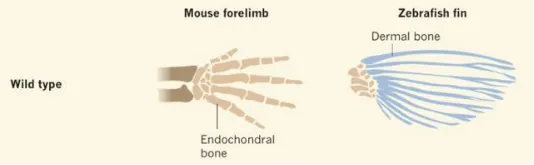
組成鰭條的膜骨,本質是一種結締組織,與軟骨內成骨的指/趾骨並非同源結構。
所以,指/趾頭不可能由鰭條發育而來,而是來源於鰭條基部的軟骨內成骨。也就是鰭條和肩帶/腰帶間連線的很小一部份,看起來的確很不起眼。站在4億年前來看,誰能想到這麽小小的一部份,在未來能夠演化到足足可以占到整個體重的50%呢?
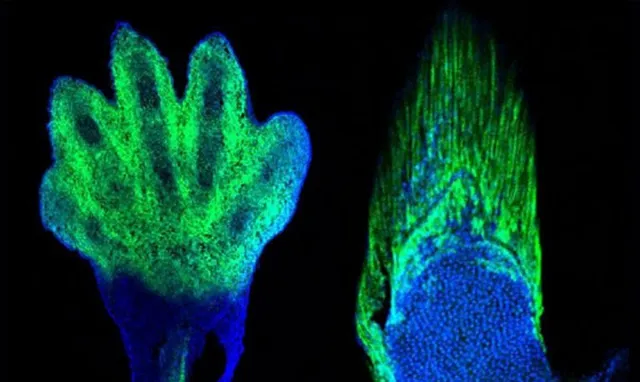
軟骨內成骨是否發展出指/趾頭,由Hox13基因家族所控制。
2016年發表於【nature】的研究發現,如果敲除斑馬魚的相關基因(hoxa13a、hoxa13b、hoxd13a),它們的鰭條就會發育得非常短。甚至隨著鰭條縮短,基部會出現了更多的軟骨內成骨,這和組成指頭的骨骼是相同的類別。相反,如果敲除小鼠的相關基因(Hoxa13、Hoxd13),它們便無法發育出手部結構。
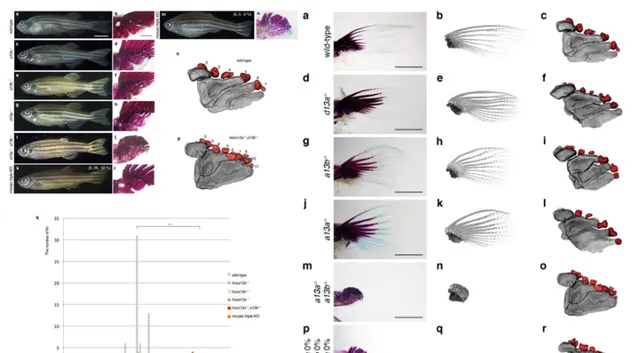
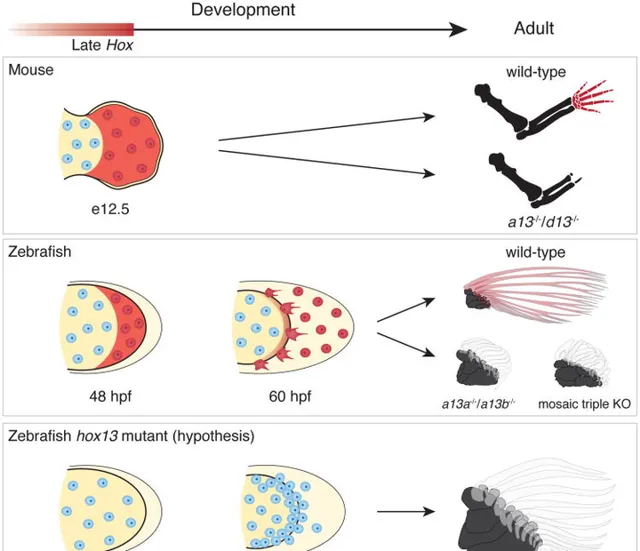
從這個實驗我們可以推測,在肉鰭魚演化的過程,大概率正是透過關閉/開啟相關的Hox13基因,來指導完成從鰭條發育到指骨發育的轉變。時至今天,偶爾出現多指癥的人類,正是與HOX13相關基因的突變有關 [5] 。
演化本身就是一場修修補補,遵守最大化利用原則。
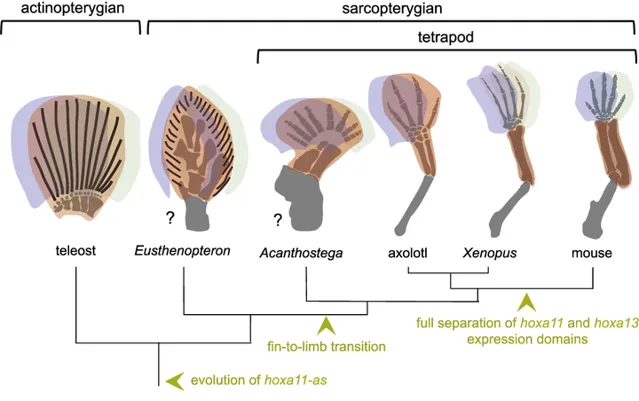
4.19億年前,夢幻鬼魚魚鰭基部的那一點點軟骨內成骨基質,經過3000萬年的時間,逐漸發展出擁有四肢骨 、 腕骨的一大類肉鰭魚,以3.9億年前的真掌鰭魚 [6] (Eusthenopteron)為代表,它們已經開始朝著陸地進軍。
然而,它們最終以失敗慘淡收場。

除了四肢還不夠強壯外,它們失敗有一個重要的原因,就是沒有演化出指/趾骨。
又過了1000萬年,3.8億年前的潘氏魚 [7] (Panderichthys)總結失敗的教訓,發展出骨化程度很高的肱骨、尺骨以及橈骨,尤其是肱骨,比同時期的其他任何魚都要長。再加上擁有強壯的肩胛骨和骨盆帶,它們可以在淺灘上進行足夠長距離的爬行。
為了適應淺灘爬行,不知經歷過多少次匍匐前行,它們終於發展出了趾骨。
雖然最開始,指/趾骨更像是一些散碎的骨頭,但透過指/趾骨支撐出更大的面積,不僅讓它們在淺灘爬行更有力量,還可以透過蹼相連,同時保持優秀的遊泳能力。
然而生物演化最忌諱的就是中間生態席位,高不成低不就,潘氏魚最終也抱憾而終。不過與它們親緣關系較近的一支卻越挫越勇,不斷改造指/趾骨,先後發展出提塔利克魚(Tiktaalik)、希望螈(Elpistostege),並最終在3.65億年前左右,演化出了具有典型指/趾骨的棘螈(Acanthostega)、魚石螈(Ichthyostega),以及圖拉螈(Tulerpeton) [8] [9] 。
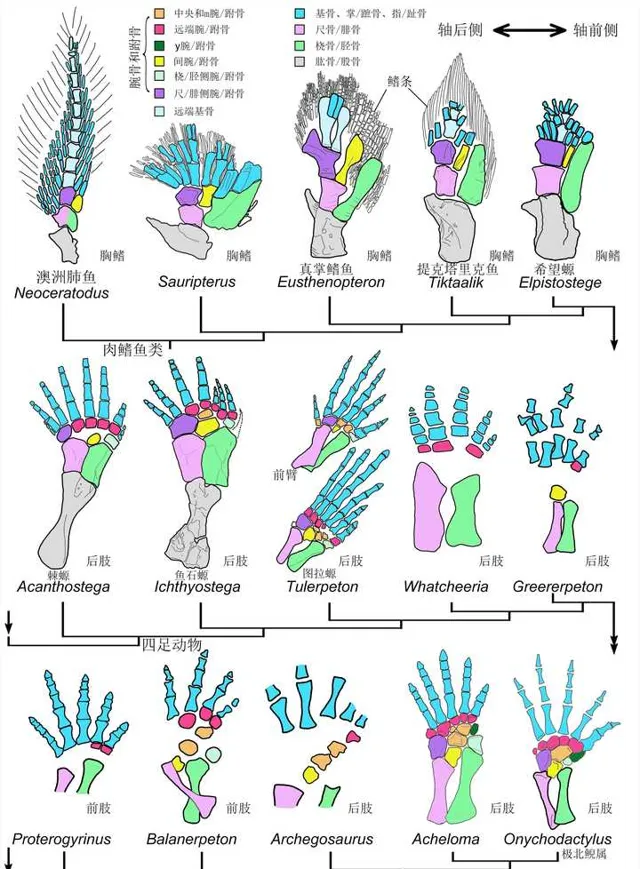
棘螈足足有著8跟指/趾骨,不過以中間六根為主。魚石螈則遺失了一根,為7跟指/趾骨。到圖拉螈時,則已經只剩下6根指/趾骨。
時間到了3.48億年前,出現了一種代表生物——彼得普斯螈(Pederpes) [10] 。

後趾5跟,前趾6跟,被認為是功能上的五趾動物。
從此以後,早期四足動物逐漸從多指型,發展為典型的五指型。
不過早期四足動物的五指型可能並不是單起源,而是多起源的。
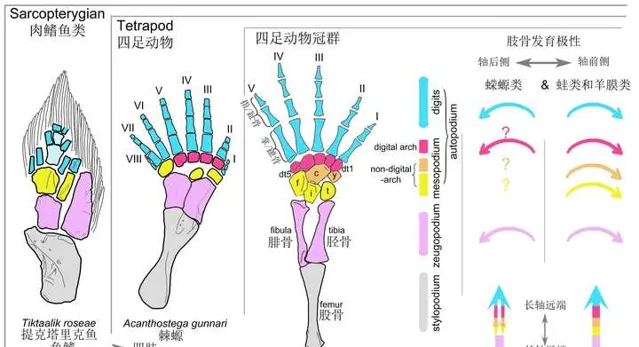
現今的所有四足動物,都是由其中一支五指型發展而來的。
其它所有指型,全都在四足動物登陸後不久,全部滅絕了。
在水環境中,魚類前進,主要面對水的阻力,最佳化身體的流體力學結構就可以了。但在登陸後,它們的身體和四肢,必須前所未有地抵抗重力。因此,無論四肢、掌骨、還是趾骨,都隨著陸地爬行產生的應力,發生全新的適應力變化。

在重力下,它們腕骨(前肢)和跗骨(後肢),發生了軸向前折。身體重量全部施加在腕骨和跗骨位置,造成骨骼發生重新排列。
手掌內部和外部兩塊小骨的內縮,使得指骨附著面積減少,最終造成了邊緣指/趾骨的退化,手指數目自然便逐漸減少。
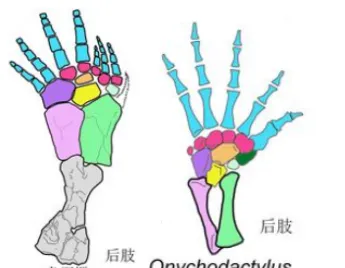
除了手指減少的第一演化壓力,來自腕部的力量支撐需要外,更少的指骨也可以避免過多手指間不必要的競爭,可使手掌上附著的肌肉更加集中,使得雙手變得更加靈活有力。因此,四足動物登陸後不久,便逐漸掌握了抓握和攀援能力,從而可以迅速輻射發展到各界各地復雜的生態席位中去,遍及山川湖泊。
你可能會有疑問,那為什麽手指沒有減少到4根以下,而是保持在5根的數目呢?
主要原因在於早期四足動物大多都是兩棲動物,它們同時還有著遊泳需求。很明顯5根手指相起比更少的手指,可以支撐起更大的面積,從而有利於遊泳。另外, 它們的骨骼的強度較低,支撐身體的指骨也不能太少。
雖然五根手指是四足動物比較原始的特征,但卻具有朝著各個方向發展的非凡潛力。
生物演化的魅力在於,原始並不等於失敗。失敗的唯一標準,是被大自然從地球生態中徹底抹除。
隨著脊椎動物向大陸全面征程,原始的五指/趾,終將隨著一次次王朝興衰,發生全新的驚人變化。
陸生脊椎動物行進的方式主要分成跖行、趾行,以及蹄行。
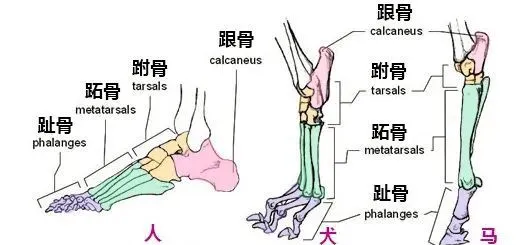
最早的四足動物全部都是跖行,至少長達1億年的時間,步態都沒有發生巨大的變化。最主要原因在於它們基本都是冷血動物,新陳代謝能力有限,它們要麽奔跑速度不夠快,要麽不能長時間持續奔跑。
時間到了2.5億年前的二疊紀末期,隨著中溫動物乃至恒溫動物的出現,動物的活動能力增加。不僅速度慢的植食動物會被吃掉,速度慢的掠食者也會被餓死。
有跑步經驗的人應該知道,高速奔跑時,擡起腳後跟,腳趾抓地,接近趾行的狀態,能奔跑得更快。隨著速度演化越來越內卷,不能保持高速步態的動物滅絕,最終活下來的步態發生了徹底改變,成了完全的趾行動物。一開始的趾行動物,也都全部具有五根指/趾頭。
不過指/趾頭越多,意味著力量更加的分散,對奔跑速度也越有影像。
因此,隨著速度需求的增強,指/趾頭就會具有退化趨勢。不過對於有著抓握和攀爬需求的動物來說,又具有反向的演化壓力來對抗趾頭的減少。
例如,今天的貓科動物,前爪還具有抓握和攀爬需求,因此保留了五爪。後肢抓握需求降低,更加傾向於奔跑,因此只有四趾。
不過最早出現趾頭減少的動物,並不是哺乳動物,而是恐龍。
二疊紀末期大滅絕事件之後,隨著合弓綱動物的衰退,主龍類崛起。從2.3億年前的三疊紀晚期開始,恐龍足足統治了地球一億多年,它們內部也是風起雲湧,王朝更替,期間不同分支發生了不同步態的變化。除了牢牢占據巨型生態席位的原始分化類群——蜥腳類是五趾外,後來的類群或多或少都經歷了高度的前進演化競爭,大多數恐龍都退化了腳趾,成了四趾動物,也有的成了三趾動物,甚至有的只有兩趾,以恐爪龍為代表。
當然由於中晚期恐龍前肢需求大大降低,嚴重退化,甚至出現了一根手指的單指臨河爪龍。

指/趾頭的變化,體現了一個物種的生存競爭壓力。它們不同形態的指/趾形,折射出的是它們曾經的輝煌。今天的鳥類依舊存在不同的趾形,同樣與奔跑、抓握等不同生境需求息息相關。
我們的祖先,合弓綱這一支,經歷二疊紀末期的大滅絕,三疊紀的大幹旱的,最終只有一些小型動物演化成了哺乳動物。為了躲避恐龍的鋒芒,它們鉆進叢林裏成為夜行動物,才博得了一線生機。
生態席位的長期穩定,也使得它們的演化異常緩慢。
最早的哺乳形態類動物出現於2.25億年前,以隱王獸為代表,幾乎和恐龍同時出現。然而一直經歷了近6000萬年的時間,到1.6億年前,它們才發展出了最早的胎生真獸類動物——中華侏羅獸。而真獸則在接下來長達近1億年的時間裏,無論體型還是生態席位都沒有發生多大的變化。

在夜行叢林生態席位上,我們祖先不僅有攀爬的需求,挖洞的需求,也有捕捉昆蟲的需求,采摘果實需求,這些都我們的雙手一直保持著保守的原始特征——五指狀態。
6500萬年前,恐龍滅絕後,整片大陸再次留下巨大的生態席位真空。
不同的哺乳動物適應不同的生態席位,再次發生了一場場特殊的前進演化競爭,四肢再次發生了巨大的變化。尤其以有蹄類這一支最為顯著,不僅演化成了今天偶蹄目和奇蹄目,一支甚至索性再次下海,演化出了鯨類,完全退化了後肢。
然而,我們祖先靈長類這一支,任它幾經滄海桑田,承擔多少興衰榮辱,終究沒有選擇退化五指,而是爬上樹梢,不斷強化手指的抓握能力和靈活性,隨後又走出叢林,不斷錘煉我們的雙手。使它成為地球上最強的攻擊性器官,最終締造了屬於我們自己的輝煌。
當然,這又是下一個故事了。從某種意義上來說,正是我們祖先把原始的五指特征堅守了3.5億年的時間,才讓我們今天有機會創造出人類王朝。
參考
- ^ Zhu, Min, et al. "The oldest articulated osteichthyan reveals mosaic gnathostome characters." Nature 458.7237 (2009): 469-474.
- ^ Kherdjemil, Yacine, et al. "Evolution of Hoxa11 regulation in vertebrates is linked to the pentadactyl state." Nature 539.7627 (2016): 89-92.
- ^ Nakamura, Tetsuya, et al. "Digits and fin rays share common developmental histories."Nature537.7619 (2016): 225-228.
- ^ Woltering, Joost M., Michaela Holzem, and Axel Meyer. "Lissamphibian limbs and the origins of tetrapod hox domains." Developmental biology 456.2 (2019): 138-144.
- ^ Malik, S., and K‐H. Grzeschik. "Synpolydactyly: clinical and molecular advances." Clinical genetics 73.2 (2008): 113-120.
- ^ Laurin, Michel, et al. "A microanatomical and histological study of the paired fin skeleton of the Devonian sarcopterygian Eusthenopteron foordi." Journal of Paleontology 81.1 (2007): 143-153.
- ^ Boisvert, Catherine A. "The pelvic fin and girdle of Panderichthys and the origin of tetrapod locomotion." Nature 438.7071 (2005): 1145-1147.
- ^ LEBEDEV, OLEG A., and MICHAEL I. COATES. "The postcranial skeleton of the Devonian tetrapod Tulerpeton curtum Lebedev." Zoological Journal of the Linnean Society 114.3 (1995): 307-348.
- ^ Ahlberg, Per Erik, and Jennifer A. Clack. "Lower jaws, lower tetrapods–a review based on the Devonian genus Acanthostega." Earth and Environmental Science Transactions of The Royal Society of Edinburgh 89.1 (1998): 11-46.
- ^ Clack, J. A., and S. M. Finney. "Pederpes finneyae, an articulated tetrapod from the Tournaisian of Western Scotland." Journal of Systematic Palaeontology 2.4 (2005): 311-346.











